Et tunc nulla erat X 10
(And Once There
Was)
Dinos: Part 3 of
3
The
Theropod March to Bird:
In the following theropod groups it
becomes increasingly difficult in determining what is deemed a dinosaur or a
bird.
Avialae: The other
major clade subdivision of eumaniraptorans is, Aviale (A-vee-ale-e) as the sister group of Deinonychosauria. The term is Latin meaning ‘bird wings’ and for
sure it represents dinosaurs that took flight leading eventually to birds. With
a temporal range from the Late Jurassic to the present 165-0 mya, avialans
include all the so-called ‘proto-bird’ ancestral dinosaurs directly linked to
birds. As an apomorphy (dependent on physical features) based clad, the general
term for Avialae is: all dinosaurs
that had feathered wings, which entails long forelimbs and fingers used for
flapping flight, along with birds that descended from them.
 |
| Artist: Rainbowleo Avialae |
 |
| Theropodal/Bird crouching |
As a group, avialans shared traits in
having elongated digits (fingers), the skeletal muscles became larger due to
hypergenesis/hyperplasia (increase in amount of organic tissue with cell
proliferation as resultant) in serving against hypothermia in particular in
hatchlings and evolved adaptations for flight or swimming due to vestigiality
as consequential to bipedalism.
One interesting note here, some
paleontologists like the esteemed, Gregory S. Paul feels dromaeosaurids may
have actually devolved from primitive avialans losing the power of flight while
keeping their feathers. This is similar to the modern ostrich and other ratites
and the time scale fits as primitive avialans came first 22 million year
earlier than dromaeosaurids. In fact, there have been dinosaur finds that have
been very confusing, like Serikornis
or Anchiornis in whether they were
dromaeosaurids or avialans.
 |
| Artist: Herschel Hoffmeyer Dakotaraptors battling T. rex |
Euavailae
(U-a-vee-ale-e), meaning, ‘true winged birds’ are avialan dinosaurs getting
closer on the march to birds. Avebrevicauda
(Hav-brev-e-caw-dah), meaning ‘birds with short tails’ in reference to the
shortened pygostyle, has no more than ten caudal vertebrae. Pygostylia (Pay-gus-style-e-uh) are
avebrevicaudan dinosaurs with short fused caudal vertebrae.
 |
| Avebrevicauda Clade |
There are only three avialan genera and
one family that are not in the direct common ancestry to birds. The genera are:
Archaeopteryx (R-key-op-teh-rix) from
the Late Jurassic 150.8-148.5 mya; Jeholornis
(Jeh-o-lor-nis) from the Late Cretaceous 122-120 mya and Rahonavis (Rah-hun-ah-vees) from the Late Cretaceous 70 mya. The
family is Anchiornithidae
(Ang-key-or-nith-ah-day) from the Late Jurassic to the Early Cretaceous 165-122
mya.
 |
| Archaeopteryx fossil |
Nonetheless that doesn’t take away from
the importance of the Archaeopteryx
discoveries. It is a transitional form of non-avian maniraptorans and birds,
but is more related to maniraptorans, like Dromaeosaurus
than it is to birds.
At 50cm/20in long, Archaeopteryx still was fully feathered with the caudal (tail) and
wing feathers rounding off at the distal ends. One fossil finding under
electron microscopy showed the feathers still retained the pigment, melanin
concluding that the feathering was black. Also, in being bird-like, Archaeopteryx’ inner ear was more like
modern birds than maniraptoran ears and the brain area dedicated to vision took
up 1/3 of brain size. However, unlike birds it still possessed a full set of
small but sharp teeth, a long bony caudal (tail), gastralia (belly ribs) and
three wing claws functional enough to grasp prey.
 |
| Artist: Paul D. Stewart Archaeopteryx |
With a length of 0.8m/2.6ft, Jeholornis was the largest avialan until
the latter half of the Late Cretaceous in which the ornithurine, Hesperornithes reached a length of
1.5m/4.9ft. There are three species in: J.
prima, J. palmapenis and J. curvipes.
Fossil remains have been found from the Chinese, ‘Jiufotang’ and ‘Yixian’
formations detailing that Jeholornis
was an herbivore subsisting primarily on cycad and ginkgo seeds along with
small fruits as witnessed in caprolithes (fossilized fruit). With seeds and
caprolithes found in the crop, gastroliths were also found in the digestive tract.
 |
| Jeholornis fossil |
Even though the shoulder blades were more on the sides of the body than on top of the back like birds, the forelimbs were larger than the hind limbs in Jeholornis and were more equipped for powered flight than Archaeopteryx was. The flight feathers were asymmetrical and aerodynamic. With a partially reversed first toe, the claw curvature alludes to the fact that it could perch on limbs. The halluces (singular: hallux ~ innermost digit of vertebrates’ hind foot) were short and along with short hind limbs, it was not a fast runner. The skull and snout were short, but robust, with J. prima lacking teeth in the upper jaw (maxilla) with three small teeth in the mandible (lower jaw), while J. palmapenis had teeth in the middle of the maxilla.
 |
| Artist: Aijuan Shi Jeholornis |
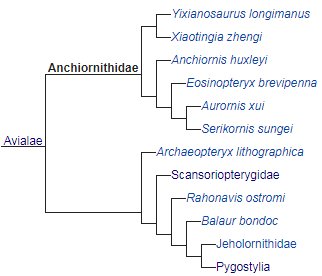
The family, Anchiornithidae had a temporal range of 165-122 mya during
the Late Jurassic to Early Cretaceous. Anchiornithids may be the most derived
and closest transitional form of avialans to the most basal of birds. Lightly built
fully feathered carnivorous/insectivorous avialans, with long legs, arms,
fingers and tails, anchiornithids anatomically resembled Archaeopteryx and even the dromaeosaurid, Microraptor, but physically on outside appearances looked like
birds. At only 30cm/12in in total length, Eosinopteryx
was the smallest avialan, while Pedopenna
and Yixianosaurus were both the
largest at 1m/3.3ft.
 |
| Anchiornithidae Clade |
The distinguishing features were in
having ‘fan-shaped’ posterior neural spines; the acromion margin (bony process
on the scapula) hooked outwards and the inside surface of the proximal part of
the fibula was flat. Also, the presence of
furrows on both sides of the phalanges (fingers) is a feature shared by all
anchiornithids. Sharing a trait with
most troodontids, the anterior dentary teeth were numerous and more closely
spaced than teeth in the middle of the tooth row.
Anchiornithidae family members
came from what is now the ‘Tiaojishan Formation’ of China and the country, Germany.
The family has nine genera, which are: Serikornis
(Say-ree-kor-niss) from the Middle-Late Jurassic 165-162 mya; Pedopenna (Puh-dop-in-nuh) from the end
of the Middle Jurassic 164 mya; Caihong
(K-hawn) from the Late Jurassic 161 mya; Anchiornis
(Ang-key-or-niss) from the Late Jurassic 160.89-160.25 mya, of whom the family
is named after; Xiaotingia
(Zhau-tin-gee-ah) from the Late Jurassic 160 mya; Aurornis (Or-ror-niss) from the Late Jurassic 160 mya; Eosinopteryx (E-oh-sye-nop-ter-ix) from
the Late Jurassic 160 mya; Ostromia
(Oss-trom-ee-uh) from the Late Jurassic 150.25 mya, in which its Haarlem TM
6428/29 fossil was once considered an Archaeopteryx
species and Yixianosaurus
(Yee-see-en-o-sawr-us) from the Early Cretaceous 122 mya.
 |
| Artist: Emily Willoughby Serikornis |
 |
| Serikornis fossil |
 |
| Artist: Zhao Chuang Caihong |
The preserved melanosomes’ cell pigments
detail splashes of orange, lite green, burnt red and blue around the cranial
and neck areas that match the iridescence of hummingbird feathers. Aside from
the iridescent feathers, Caihong is
also the earliest known not just dinosaur, but animal to support asymmetrical
feathering. Although asymmetrical feathering is on modern birds’ wings, they
were on Caihong’s tail. Only the body
feathering was shaggy and not iridescent.
 |
| Caihong feather fossil |
Eosinopteryx, is the
smallest of anchiornithids, with its fossil remains coming from the western
portion of China’s, ‘Tiaojishan Formation’. the remains were well preserved in
presenting virtually a wholly articulated skeletal anatomy of a young adult
with feather impressions.
 |
| Eosinopteryx fossil |
 |
| Artist: Emily Willoughby Eosinopteryx |
 |
| Anchiornis fossil |
 |
| Anchiornis wing fossil with skin (white) |
 |
| Artist: Zhao Chuang Anchiornis |
Xiaotingia as well came
from the ‘Tiaojishan Formation’ of China’s, Jianchang County of Liaoning
Province. With a length of 0.6m/2ft, it had long and robust forearms, which at
first was thought to support powered flight. Due to this thinking, it was
proclaimed to be the first bird, trumping Archeopteryx
of that crown by 10 million years. But as it stands, both cannot claim the
crown, for neither one qualifies to being a bird.
 |
| Artists: Xing lida & Liu Yi Xiaotingia |
 |
| Xiaotingia fossil |
Euavialae and Avebrevicauda
are two avialan subclades that led to Pygostylia. I won’t go much into
detail with these two as they are simply a sequential line to pygostylians. All
three have a temporal range of 131-0 mya with a chronological order in
phylogenetically putting the most primitive first, which would be: euavialans
=› avebrevicaudans =› pygostylians.
Euavialae literally
means, ‘true winged birds’ and is the major separation between feathered
dinosaurs and modern birds. Besides in the clade Euavialae leading to birds through Avebrevicauda, it holds one genus that was monotypic as a euavialan
in Jixiangornis. Many scientists call
euavialans ‘primitive birds’, but as a scientist and junior paleontologist
myself, I’m still going to refer to them as dinosaurs. Under ‘Birds’ I’ll give the true definition of
birds with a list of anatomical traits.
 |
| Artist: Jack Wood Jixiangornis |
 |
| Jixiangornis fossil |
 |
| Avebrevicauda Clade |
Avebrevicauda literally means, ‘birds with short tails’ and so named to separate dinosaurs from their reptilian long tailed ancestors to those with ten or fewer free caudal (tail) vertebrae. Considered a stem based clade, Avebrevicauda contains all known avialans that had lost the hyperextended pedal/pes (foot/feet) digit II (second toe). This is a form of ‘oligodactyly’ when a digit from a limb never develops and is much more common than ‘polydactyly’ when an extra digit (toe or finger) is developed. Both anomalies are caused from genetically or familial altercations in the fetal stage. The Avebrevicauda clade includes the family, Omnivoropterygidae (Alm-nih-vor-op-teh-ridge-ah-day) and the whole of Pygostylia descendants, that includes the clade node order, Confuciusornithiformes (Cun-fusch-suh-sor-ah-nith-e-for-meez) and its family and the node based clade, Ornithothoraces along with its subdivisions.
Omnivoropterygidae: With a temporal range of
125-120 mya during the Early Cretaceous, omnivoropterygids were primitive
avebrevicaudans and most likely were a short and regional occurrence with all
coming from a 120 million year old section of the ‘Jiufotang Formation’ of
Chaoyang, China. Because there are only teeth in the upper premaxillaries,
omnivoropterygids are considered omnivores, hence the name. There are two
omnivoropterygid genera with one species in each and they are: Omnivoropteryx (Alm-nee-vor-op-teh-rix)
and Sapeornis (Sape-or-niss). There
is an order above Omnivoropterygidae, but there are no members
outside of the family Omnivoropterygidae; its name is, Omnivoropterygiformes. Also, due to some
similarities, although there are some primitive differences with Omnivoropteryx to oviraptorosaurs, those
similarities may make Omnivoropteryx
a junior synonym of Sapeornis and the
name may be abandoned which would also replace the family name of Omnivoropterygidae to Sapeornithidae.
Just keep that in mind for future referencing. Whew…a lot of Omni’s in
there.
 |
| Artist: Yike Xu Sapeornis |
 |
| Sapeornis fossil |
Pygostylia: Pygostylians
occurred, 131-0 mya from the Early Cretaceous to Present. Pygostylian features
closed the gap in what is a dinosaur and what is a bird. In fact, many esteemed
paleontologists indeed do refer to them as primitive birds. However in my
perspective, perhaps proto-birds, but not birds, at least not in saying they
possess all the same characteristics as Neornithes
(modern birds).
 |
| Bird anatomy with pygostyle |
Aside from the pygostyle, there’s
considered three other main features of pygostylians and they are: the total
absence of an accessory joint found in most archosauromorphs known as the
‘hyposphene-hypantrum’, a reversed pubic bone and a bulbous medial condyle
(round prominence at the end of a bone) of the lower leg bone known as the
tibiotarsus.
 |
| Pygostylia Cladogram |
Confuciusornithiformes
(Cun-few-shus-or-nith-e-for-meez) contains members that first evolved the
bird-like beak occurring 131-120 mya in the Early Cretaceous. Also, according
to the fossil record, it appears that confuciusornithiforms are the first to
evolve vitellogenesis, commonly called, ‘yolk deposition’ where nutrients being
deposited in the oocyte, or female germ cell involved in reproduction of
lecithotrophic (oviparous animals that lay eggs with little or no other
embryonic development within the mother) organisms.
All known confuciusornithiform fossils
lacked teeth in having a beak instead of a snout, but this was independent of
birds. The beak tip ended in a point. Generally, they were very small but with
robust forelimbs supporting long and narrow pennaceous feathers.
The order, Confuciusornithiformes is divided into the family, Confuciusornithidae
(Cun-few-shus-or-nith-ah-day) and the most primitive confuciusornithiform, in
the monotypic genus, Eoconfuciusornis
(E-oh-cun-few-shus-or-nees).
 |
| Artist: Nobu Tamura Eoconfuciusornis |
 |
| Eoconfuciusornis fossil |
 |
| Artist: Jaime Chirinos Confuciusornis feducciai |
For the genus, Confuciusornis there are currently three confirmed species and they
are: C. sanctus (sanque-tuce), C.
dui (d-u-i) and C. feducciai
(feh-dook-say). Confuciusornis
species lived 125-120 mya ago during the Early Cretaceous. There have been hundreds of well preserved
and articulated fossils found in the ‘Yixian and Jiufotang’ formations of
China. In fact, on a 100 meter squared area forty fossils were found in
articulated condition. This was a result of volcanism as all the fossils were
found in tuff sediment, which is the result of compacted volcanic ash.
 |
| Confuciusornis morphology |
 |
| Confuciusornis fossil toe scales |
 |
| Confuciusornis |
 |
| Cofuciusornis wingbone |
As far as flying is concerned, Confuciusornis could fly; how well is
still up for debate. The unusually long forelimbs supported primary
asymmetrical flight feathers, its fused wrists and ossified sternum with a
central keel, possessing strut-like coracoid bones which could form a solid
base for wing muscle attachments and its deltopectoral crest which provides
powerful upstrokes all point to the fact it could fly. However, it lacked an
aula (an extension process formed by feathers anchored to the first digit of
the hand as in modern birds), the forearms lacked any quill knobs and the heavily-built
and immobile skull was incapable of the kinesis of modern birds in the ability
to raise the snout relative to the back of the skull.
 |
| 1st: Franz Anthony C. sanctus 2nd: Daniel Bensen C. dui |
 |
| Artist: Matthew Martyniuk Changchengornis |
Changchengornis lived 124.6 mya
during the Early Cretaceous although closely related to Confuciusornis, it was much smaller at 20cm/8in long. It is a
holotype with the fossil remains coming from the ‘Chaomidianzi Formation’,
known previously as the lower section of the ‘Yixian Formation’ of china was
very much like the Confuciusornis
species in physiology and anatomical traits. As in Confuciusornis, the male had the two ribbon-like tail feathers, the
foot structure was adapted for perching; it possessed the same three fingered
claws and the beak was toothless and both had a long pygostyle.
 |
| Changchengornis fossil |
Ornithothoraces ~ One way to
look at Ornithothoraces
(Or-nith-o-thor-ace-eez) is that when it comes to evolution, this group is kind
of like the halfway house between dinosaurs and birds. Technically, this means
nothing but perception and perception is not always reality, for a ‘halfway’
perceived evolved species leads to teleology as if one particular taxon group
is ‘predestined’ to evolve into another taxon. This halfway house rule of thumb
is not scientific in application, however, in turning the page towards
descending speciation it does give the layman, as myself, a clearer picture of
relationships.
 |
| Ornithothoraces Cladogram |
Ornithothoracines’ skeletal anatomy
included a keeled sternum (breastbone), elongated coracoids (hook-like
structures on lateral edge of scapula), the shoulder’s glenoid joint and
semi-rigid ribcage had been modified from earlier avialans. Also, there was an
absence of gastralia (Singular: gastralium dermal bones found in ventral body
wall of reptiles), the scapula terminated distally into a sharp point and the
pygostyle, although primitive, fused the most distal caudals into a short tail.
In various literature and even a few
early periodicals, you might come across the name, enantiornithean as,
‘enantiornithine’ and that also goes for euornithean as, ‘euornithine’. This
infers that they are subfamilies but they are not. So, the ‘ine’ suffix is
wrong.
Enantiornithes
(E-nan-tee-or-nah-theez) ~ Enantiornitheans are the most speciose avialans of
the Mesozoic found around the globe, except Antarctica and were widespread in
the Early Cretaceous Asia and Europe coexisting with relatively modern birds
for nearly 24 million years until the Cretaceous/Paleogene Extinction. All of
them possessed teeth and clawed fingers and besides the teeth and claws, the
main difference between enantiornitheans and birds was in the articulation
between the scapula and coracoid. This entails the shoulder bones where the
concave/convex socket joint involving the scapula and coracoid in
enantiornitheans operated exactly the opposite or in reverse as do modern
birds.
Evidence shows that there wasn’t much
parental care concerning enantiornitheans, much unlike modern birds’ devoted
parental care. With enantiornithean hatchlings hatching from eggs laid on the
ground, they would soon have to scamper up trees before being eaten by other
animals. Within a week they had feathers and we know this from observing the
many hatchlings that were encased in tree sap that fossilized into amber. These
amber fossil finds show exquisite detail of feathers, skin and scale features
such as morphologies and coloration.
 |
| Enanriornithean chick encased in amber |
Before getting into enantiornithes
families, first off we’ll touch base on the holotypic basal enantiornitheans.
There are at least 52 species of enantiornitheans, but we’ll start with nine
holotypic genera in: Protopteryx
(Pro-top-tuh-rix) from the Early Creataceous 131 mya; Iberomesornis
(I-beh-roe-matsor-neez) and Eoalulavis
(E-o-ah-lu-lay-viss) both from the Early Cretaceous125 mya; Liaoningornis (Leh-ow-ning-or-neez) and Eoenantiornis (E-o-en-an-tee-or-neez)
both from the Early Cretaceous 124.6 mya;
Sinornis (Sign-or-neez), Cathayornis (Kuh-they-or-neez), Eocathayornis (E-o-kuh-they-or-neez)
with all three from the Early Cretaceous 120 mya and Flexomornis (Flex-o-mor-neez) from the Late Cretaceous 96 mya. Even
though Cathayornis and Eocathayornis have similar names, they
are only similarly related. Sometimes these holotypes are grouped into a clade
as, Euenantiornithes.
 |
| Artist: Goombasaurusrex Protopteryx |
 |
| Protopteryx fossil |
 |
| A. Protopteryx B. Eoenantiornis |
 |
| Sculptors: José Manuel & Benito Álvarez Iberomesornis |
It was small, around the size of a house
sparrow with a length of ~16cm/6.3in. With short wings and a wingspan, Iberomesornis possessed a claw on each
wing. Equipped with superb flight agilities and perching ability in utilizing
curved toe claws, it most likely caught small insects on the wing to swallow in
flight and/or small ground invertebrates to eat while perched.
 |
| Sculptor: Jose Salas Eoalulavis |
 |
| Artist: Jack Wood Lianoningornis |
 |
| Artist: Jack Wood Eoenantiornis |
 |
| Eoenantiornis fossil |
 |
| Artist: Danneart Sinornis |
 |
| Sinornis fossil |
 |
| Artist: Leau Bellon Cathayornis |
 |
| Artist: Scott Reid Eocathayornis |
Eocathayornis skull traits
show the premaxilla had a long slender nasal process. There were four pointed
small teeth anchored in the premaxilla. Each tooth had a constriction at the
crown’s base, which is typical of all toothed enantiornitheans. As in Archaeopteryx and Cathayornis, the cranial tip of the premaxilla is hooked and
edentulous (toothless). The maxilla’s pointed dorsal process was caudally
inclined.
Claws were present on the three digits
(fingers). The relatively short sternum had a pair of long caudo-lateral
processes and a low and caudally distributed keel. Even though this was a
primitive enantiornithean, traits of the scapula were advanced and along with
the forelimb and feathering wing structure, Eocathayornis
was quite capable of powerful flapping for flight.
 |
| Artist: Jack Wood Flexomornis |
The name, Flexomornis is in reference to its unique flexed shoulders. In all
appearances it looked like a modern bird, but still retained teeth, forelimb
distal phalanges (fingers) and claws. As evidenced by the diagnostic scapula,
it had a slight variance in muscle attachments than the older enantiornitheans.
This would infer that it was a large enantiornithean at ~ 33cm/13in from head
to tail tip.
 |
| Artist:Jack Wood Gobipteryx |
Around the size of a current partridge
at 30-33cm/11.8-13in, Gobipteryx,
which occurred 72 mya, had a toothless beak formed by the fusion of premaxillae
bones. The scapula was long and was well suited to anchoring flight muscles. In
addition, the scapula contained a prominent glenoid labrum (a
fibrocartilaginous structure that rims the cavity of the shoulder blade) that
tapered backwards ending in thin rods.
 |
| Gobipteryx fossilized eggs |
Pengornithidae: Pengornithids
come from China’s, ‘Huajiying Formation’ and the ‘Jiufotang Formation’. These
were one of the most primitive groups of enantiornitheans with numerous
mandibular (lower jaw) small teeth and on the other end possessed stout legs.
With a temporal range in the Early Cretaceous of 130.7 mya, all pengornithid
fossil species have come from the ‘Huajiying’ and ‘Jiufotang’ formations of
China’s Liaoning and Hebei provinces.
Along with a shortened tail ending in a
rounded pygostyle, pengornithids’ main skeletal feature was a hooked outgrowth
of the shoulder blade. Most all, enantiornitheans had four long backward
projections coming from their breastbones; pengornithids only had two.
Pengornithids retained some primitive traits in an elongated fibula and
retaining metatarsal V (5th toe).
Pengornithids had the ability to fly,
but the feature noted best by them is their capability to vertically climb.
Stiff tail feathers, as in woodpeckers were present acting as a prop while the
feet were adapted to hold onto branches with the claws well enforced to anchor
into bark. With these features pengornithids were predominately insectivores
after insects, grubs and other invertebrates hidden amongst the bark; although,
Eopengornis is suspected of also
being more of a ground dwelling carnivore, due to its recurved teeth. Most were
also aerobatic flyers as evidenced in the short fan tail of the Chiappeavis fossil.
Pengornithidae: this is the most basal family of enantiornitheans consisting of four genera; they are: Eopengornis (E-o-pen-gor-neez) from 130 mya in the Early Cretaceous; Pengornis (Pen-gor-niss), Parapengornis (Par-ah-pen-gor-neez) and Chiappeavis (Chi-ap-pay-viss) all three from 120 mya in the Early Cretaceous.
Pengornithidae: this is the most basal family of enantiornitheans consisting of four genera; they are: Eopengornis (E-o-pen-gor-neez) from 130 mya in the Early Cretaceous; Pengornis (Pen-gor-niss), Parapengornis (Par-ah-pen-gor-neez) and Chiappeavis (Chi-ap-pay-viss) all three from 120 mya in the Early Cretaceous.
 |
| Artist: Jack Wood Parapengornis |
 |
| Parapengornis fossil |
Until the discovery of Parapengornis with published results in
June, 2015, Pengornis was the largest
enantiornithean at 25cm/9.8in. But only for a brief few months as its results
were published through the internet in January, 2015. Pengornis fossil remains come from the ‘Jiufotang Formation’ of
Dapingfang, Liaoning province, China.
 |
| Artist: Jack Wood Pengornis |
 |
| Pengornis fossil |
 |
| Bohairnithidae Clade |
Bohaiornithids possessed long and thin
lateral trabeculae (bony projections on posterior end of sternum) that extended
backwards and outwards. The scapulae (shoulder blades) slightly curved
downwards creating a convex top edge and lower concave edge. Digit II, the
second innermost toe of the pes (feet) were the thickest, while digit III as
the middle toe was the longest. Male bohaiornithids also possessed the two tail
ribbon feathers that were iridescent in life.
There are currently eight known genera
of bohaiornithids that all came from Early Cretaceous China and they were: Zhouornis (Zow-or-neez) from ~125.4-120
mya during the Early Cretaceous; Sulcavis
(Sul-cay-viss) from 125-121 mya during the Early Cretaceous; Shenqiornis (Shen-key-or-neez) from 122
mya during the Early Cretaceous; Longusunguis
(Lon-gus-un-giss) and Parabohaiornis
(Par-ah-bow-hay-or-neez) both from 120.3 mya during the Early Cretaceous and Bohaiornis (Bow-hay-or-neez), Fortungauvis (For-toon-gau-viss) and Linyiornis (Lin-yee-or-neez) with all
three from ~ 120 mya during the Early Cretaceous.
 |
| Artist: Stephanie Abramowicz Sulcavis |
 |
| Sulcavis fossil skull |
Other anatomical characteristics of Sulcavis were: the skull was longer than
the cervicals; the posterior portion of mandible was set deeper than anterior
end; posterior portion of quadrate (squarish bone which articulates the jaws) was
straight; the retroarticular (cartilage bone that forms at the posteroventral
tip of Meckel’s cartilage where the interoperculomandibular ligament attaches) directionally
descends and pes metatarsals (foot bones) II and III aligned with I.
 |
| Artist: Jack Wood ♂ Bohaiornis |
At ~ 47cm/18.5in, Bohaiornis was one of the largest bohaiornithids. Its dentary
(upper jaw bone) was straight and half the length of the mandible (lower
jawbone). Another couple of unique traits were that the sternum was markedly
wider than long while the acrimon (process on the outer end of the scapula
extending over the shoulder joint) is very robust. The nasal cavities were also
very broad.
 |
| Artist: Eloy Manzanero ♀ Bohaiornis |
One fossil specimen had small rounded
pebbles in its thoracic region. Along with its cranial and dental morphology,
and the number and distribution of teeth, Bohaiornis
most likely lived a raptorial ecology feeding on small vertebrates and large
arthropods like spiders and insects.
Longipterygidae: With a 125-110
mya span of known longipterygid species, they’ve only been found in the ‘Jiufotang’ and ‘Yixian’ formations of China. Where pygostyles are usually a series
of short fused vertebrae, longipterygids had an unusually long pygostyle.
Making up over 60% of the skull, the
snout was extremely long with the tip of the snout supporting teeth in the
premaxilla (upper front jaw) and rostral portion of the dentary (lower jaw).
The shape of the snout was straight, but slightly concave just behind the
nostrils and ended in a point. The feet were some of the most specialized of
enantiornitheans where all the toes were of the same length.
Currently there are seven genera listed
under the longipterygid family clade with one species within each genus. The
genera are: Longirostravis
(Lon-gee-row-stray-vis) from ~125 mya; Shanweiniao
(Shan-way-now) from 122 mya; Boluochia
(Bwaw-lwaw-chuhr-uh) from 121.6 mya; Longipteryx
(Lon-gip-ter-ix) from 120.3 mya; Camptodontus
(Camp-toe-don-tus) from 120 mya; Shengjingornis
(Shen-jing-or-nis) from 120 mya and Rapaxavis (Rah-pax-uh-vis) from 113 mya.
All lived during the Early Cretaceous in what is now the ‘Jiufotang’ and ‘Yixian’ formations of China.
 |
| Credit: PNG Clipart Longirostravis |
The word, ‘longirostravis’ is Latin
meaning, ‘bird with a long rostrum’. The rostrum, or snout was long and
slightly curved at the tip while supporting five pairs of small conical teeth.
Most likely the snout was used for probing in mudflats to get at small
crustaceans or shellfish. The sternum was unusually shaped and was much like
moose horns with a pair of three-pronged projections. No more than 17.8-22.9cm/7-9in,
Longirostravis was no bigger than the
extant bobwhite quail.
 |
| Artist: Scott Reid Shanweiniao |
 |
| Artist: Sydney Mohr Shanweiniao |
 |
| Artist: Daniel Bensen Boluochia |
 |
| Credit: Nix Illustration Longipteryx |
Longipteryx is the dinosaur
theropod equivalent of a modern day kingfisher as it was a well-equipped
aquatic feeder to dive, nab small fish or crustaceans, then fly back up to
perch on a tree and consume its meal.
The enantiornithean, Camptodontus is not to be confused with
a beetle genus of the same name and the species name for an Astragalus plant. The term means, ‘bent
tooth’, but is currently the unofficial name for this distinct enantiornithean.
 |
| Artist: José Carlos Cortés Camptodontus |
 |
| Artist: Jack Wood Shengjingnornis |
The body length of Shengjingornis was 20cm/7.9in with a wingspan a little over the
body length at 22cm/8.7in. In contrast to the body, the legs were long distally
ending in curved claws, while each of the wings possessed three short curved
claws. The wing features, including a cake-like sternum, indicates it was a
strong flapping theropod. With a bent downward snout ending in rather large
sharp conical teeth just in front of the nasal alludes to a fact that this
longipterygid was a specialized feeder. But what it fed on is strictly speculative;
however, it is possible that it was well equipped to probe tree bark for
insects, grub worms and other invertebrates. Or, but this is really stretching
it, skim the surface of bodies of water to ensnare and trap small fish, aquatic
insects and crustaceans.
 |
| Shenjingornis fossil |
 |
| Artist: Jack Wood Rapaxavis |
 |
| Rapaxavis fossil |
The fossil remains reveal the
caudolateral processes of the quadrangular-shaped sternum possessed two
prominent branches forming a V-shape ending into one small xiphoid (lowest
division of sternum) branch. As a character of enantiornitheans, a procoracoid
is absent. Although gastralia were preserved in the proximity of the pelvic
girdle, there is no evidence of uncinate (hook shaped) processes that usually
accommodate gastralia.
One item noted in the fossil remains of Rapaxavis is that it is the only
exception within the whole clade of enantiornitheans that the derived manus
(hands) lack unguals (pertaining to claws, nails or hooves), even though it
possessed keratinous sheaths of the pedal unguals. Also, like the
enantiornithean Concornis, Rapaxavis possessed a pair of paracoracoidal
ossifications (pectoral bones). No one as yet knows the exact function of these
bones and within the entire clade of Avialae,
these are the only two species that possess this unique ossification.
 |
| Rapaxavis skull |

With a temporal range of 80 mya during
the Late Creatceous, a body length of 17cm/6.7in, a wingspan of 40cm/15.8in and
with the fossil remains found in Alabama’s ‘Mooreville Chalk Formation’, Halimornis, as in most all avisaurids,
was among the largest, most diverse and last of the enantiornitheans.
 |
| Artist: Scott Reid Halimornis |
 |
| A young hatchling enantionithean encased in amber |
Birds:
It has gotten to the point with many new
discoveries and anatomical analysis that maybe the phraseology shouldn’t be
bird-like dinosaur, but dinosaur-like bird.
 |
| Theropodal evolution to bird |
The common ancestor to modern birds
diverged from other paravians around 95 mya. Molecular biology confirms this.
Anatomy confirms this and paleontology confirms this…not only did birds
speciate from ancestral dinosaurs...birds are dinosaurs. In fact, T. rex is more closely related to the
chicken in genetics and time than it was to the earliest true dinosaurs. Gallus jungle fowl appeared 50 mya,
while T. rex reined 68-66 mya; the
first dinosaur appeared 231.4 mya. So as you see, there is only a difference of
16 million years between T. rex and
the Gallus jungle fowl, while T. rex showed up 163.4 million years
later after the first dinosaurians appeared.
Genes in birds are very much like
dinosaur genes, it’s just the sequence that varies in what genes have been
switched on and what has been switched off. Bird embryos carry an exacting
average of caudal (tail) vertebrae as dinosaurs had. As the embryo further
develops, the gene for tails is switched off with the tail vertebrae being
absorbed for building blocks somewhere else in the developing fetus. Chickens
early on in fetal development possess tiny teeth that barely erupts the gum
line, but then are switched off from further development and are absorbed. On a
regular basis, molecular biologists inject a viral component into developing
chicken embryos that maintains the switch, switching it from off to stay on,
thus further developing the embryo’s teeth. Upon hatching, the chick has a
well-stocked mouth of teeth that not only appear shaped as theropod dinosaur
teeth, but in positioning, are recurved backwards like theropods.
Female birds grow a special type of bone
in their limbs between the hard outer bone and the marrow. Called the medullary
bone, it is rich in calcium and is used up in making eggshells during breeding.
The female birds that produced it reabsorb calcium when they have finished laying
eggs in rebuilding the medullary for the next breeding season. Medullary bone
has also been found in many theropod fossils from allosaurs, tyrannosaurs and
tenontosaurs, which also aids in determining the sex of the fossil remains. The
medullary also has shown up in female subadult fossils indicating sexual
maturity was reached before adulthood, just like in humans.
The skeletal anatomy, morphology,
physiology and behavioral evidence conclusively place birds within the
evolutionary radiation of theropod dinosaurs. With respect to growth and
energetic physiology, the first birds were simply feathered dinosaurs. The
evolution of the novel bird pattern in modern forms occurred later in the
group’s history. As the result, modern birds are the most speciose amniote with
well over 10,000 species.

Although ornithischian filaments and theropod proto-feathers are not obvious in birds, dinosaurs were the first animal to support feathers. Reptilian scales evolved 320-310 mya; dinosaurs retained them and passed scaled legs over to birds.
Now, with over 10,000 extant species,
with this much diversity, birds by far outnumber extant mammals and reptiles
two to one, so let’s get to ‘em.
Euornithes (E-u-or-nah-theez)
~ is the clade leading directly to modern birds. This is the line of theropods that
became what we think of as birds. So, all feathered and toothless theropods
capable of flight and all modern birds with the exceptions of penguins and palaeognaths
(flightless ostrich-like birds) belong to this group. Therefore, it also
includes the most common recent ancestor to all modern birds. The temporal
range is from 130.7 mya to the Present.
Euornitheans are the first true birds;
in fact Euornithes means ‘true bird’.
Feathers, a short fused pygostyle, toothless beaks, clawless forelimbs, hollow
bones, wrist joints positioned for flapping wings, a bastard wing, air sacs, tridactyl
feet/hands, homeothermy, cranial kinesis, smaller size, female medullary bone, brooding
and perching all first evolved in earlier theropods to culminate into an animal
we call bird. These adaptive evolvements were originally not intended for
flight, but through natural selection and speciation came together and gave us
the bird.
 |
| Evolution of the theropod manus (hand) |
The avian respiratory system plays a
role in thermoregulation while the lungs are much smaller when compared to other
vertebrates, but are supported by nine theropod inherited air sacs that play a
big role in respiration. Due to air sacs allowing for a unidirectional flow of
fresh air into the lungs, the lungs are constantly supplied with oxygen. So as
air sacs fill, the lungs are also filling, but as air sacs and the lungs empty,
the air sacs empty into the lungs keeping them oxygenated with a constant
supply of fresh air.
 |
| Bird air sacs |
The basal euornithean holotypic genera
are, Archaeorhynchus (Are-kay-o-rin-cuss)
from 125-120 mya; Jianchangornis (Jeye-uh-chan-gor-nee-is)
from 120 mya; Zhongjianornis
(Zhone-gee-jeye-or-nis) from 120 mya and Chaoyangia (Chai-yang-nee-uh) from
120mya. The ornithuromorphan genus sister group is Schizooura (Ski-sue-u-rah)
from 120 mya. These are the most primitive euornitheans; a few even retained a
few teeth and small manus (hand) claws. But even though they are considered
proto-birds, they had more in common morphologically, physiologically,
anatomically and in physical functionality with modern birds than with Sinornis or any other enantiornithean.
All their fossils originate in China.
With the fossil remains coming from the ‘Yixian
Formation’, Archaeorhynchus’ fossils
display well preserved complete skeletons measuring 20cm/7.9in long. Even
associated with the head, neck and tail regions, feathering was preserved. It
is one of the earliest known ornithuromorph that had a toothless β-keratin beak
(proteins made from both fibrous and matrix components).
 |
| Artist: Brian Choo Archaeorhynchus |
Numerous gastroliths (stomach stones)
were found and with the amount shows that the stones were intentionally
swallowed and not incidental. This suggests that it was an herbivore.
 |
| Archaeorynchus fossil with feather pigment |
 |
| Artist: Scott Reid Jianchangornis |
 |
| Jianchangornis fossil |
 |
| Artist: Jack Wood Zhongjianornis |
 |
| Zhongjianornis fossil |
 |
| Artist: ©Daniel Benson Chaoyangia |
 |
| Chaoyangia fossil |
 |
| Artist: Jack Wood Schizooura |
 |
| Schizooura fossil |
Two other basal euornitheans have been
discovered and they are: Patagopteryx
(Pat-ah-gop-tuh-rix) and Vorona (Voe-roe-nah).
Their fossil remains were found outside China proving to be a bit unusual in
form anatomically.
 |
| Artist: Jaime Chirinos Patagopteryx following sauropods |
 |
| Patagopteryx internal anatomy |
 |
| Patagopteryx skeletal |
Patagopteryx could’ve been
an herbivore, or a carnivore, or even an omnivore as its hard beak could’ve
sufficed any diet. Also, the second toe had a curved claw, but unlike
velociraptors it wouldn’t have been useful as a weapon.
Due to the fragmentary 70 mya fossil
remains from the ‘Maevarano Formation’ in Madagascar, Vorona phylogenetic classification is difficult to determine,
but it’s clearly a basal euornithean and may later after further study, or
other fossil finds even be classified as a basal ornithuromorph.
 |
| Artist: Hyrotrioskjan Gargantuavis |
By far in being the largest euornithean,
Gargantuavis was ~ 2.2m/7.3ft in
total length and weighed in at ~ 140 kg/310 lbs. It thus far, is the largest
known bird of the Mesozoic Era. With reduced wings, it was nonvolant (incapable
of flight) as considered secondarily flightless. Even though it had a rather
broad pelvis, the hips were narrow and bird-like, still though, the girth of
the hips prevented the two ilia (singular: ilium which is the upper part of
bone forming each half of the pelvis) from meeting. The acetabulum (hip socket)
was set close to the front rather than more towards the middle of the pelvis.
Osteological studies on its remains show that it had a rapid early growth spurt
followed by an extended period (~ 10 years) of slow cyclical growth before
attaining skeletal maturity.
Sedimentary and mineralogical studies
affirm that its habitat consisted of episodes of semi aridity and seasonal
climate. This, in conjunction with living on an isolated island archipelago
land mass, natural selection evolved Gargantuavis
into unique traits. This fact, made it difficult to classify as it had no
lineage and was not related to the ratites such as the extinct moa or extant
ostrich. However, the bone studies showed an avian/euornithean nature in
osteological features.
Ornithuromorpha is the most
inclusive clade containing extant birds and was the most derived avian group
during the Early Cretaceous. Ornithuromorphans range from 130.7 mya during the
Early Cretaceous to the Present. From the primitive ornithuromorph to modern
birds, ornithuromorphan birds have a temporal range from 130 mya to the
Present.
 |
| Ornithomorpha cladogram |
Currently, there are around twenty known basal holotypic ornithuromorphs and two families with their genera. Some holotypes are: Ambiortus (Am-be-or-tus) from 130 mya; Yumenornis (U-men-or-niss) from 125 mya; Eogranivora (E-aw-gran-e-vor) from 125 mya; Changzuiornis (Chun-su-e-or-niss) from 122.1 mya; Bellulornis (Bell-ull-or-niss) from 120.5 mya; Gansus (Gan-souss) from 120 mya; Iteravis (I-tur-a-vuh) from 120 mya; Jiuquanornis (Jee-kin-or-niss) from 119 mya; Changmaornis (Chun-ma-or-niss) from ~102.5 mya; Apsaravis (App-sar-a-vuh) from 78 mya and Alamitornis (Al-uh-me-tor-niss) from 70 mya. There is some questionable research on whether the Iteravis fossil is actually another fossil specimen of, Gansus. Further research will determine the actual classification.
 |
| Artist: Nobu Tamura Eogranivora |
Initially, the fossil remains of Eogranivora was first misidentified as Hongshanornis, but after further
observations it was noted that the remains detailed longer wings, was
edentulous, had stouter shorter legs with the backward pointing digit I (commonly
known as the perching toe while technically known as the hallux) and the first
metatarsal were missing, thus having only three forward pointing toes in digits
II, III and IV. This, as compared to modern birds, suggests that it was more
terrestrial rather than arboreal. However, the legs wouldn’t have had the
efficacy to run fast, but with fairly developed wings it would’ve flown off
instead trying to outrun any danger.
 |
| Eogranivora fossil |
With the evolution of flight, Eogranivora is a direct link of Early
Cretaceous birds to modern birds and has produced associations into the trophic
habits and digestive abilities of early birds and their closest relatives to
extant birds. Information from its fossil reveals the changes in the digestive
system that occurred with the evolution of flight, through the structural
changes of a lighter digestive system.
 |
| Artist: Mark A. Klingler Gansus |
Although the wings were slender and
incapable of vigorous flight, they still retained flight feathers and were just
long enough with a stout sternum to be volant (capable of flight), but short
enough to be held back against the body during dives.
With elongated manus (hands), the
wing-bones of Gansus were thin,
indicating with unstiffened wings, a more gracile flight pattern rather than a
strong burst of flapping. The pedes (feet) had interdigital webbing, making it
one of the first birds to utilize more efficient aquatic propulsion.
 |
| Gansus fossil |

Earlier, we mentioned the comparison between Gansus and Itreavis fossils and how initially researchers first thought they were one and the same species. In comparing the above comparison between one of the Gansus fossils and the fossil of Itreavis, you can see how at first it was misidentified.
 |
| Artist: Scott Reid Ambiortus |
At 40.1cm/15.8in, including the tail, Ambiortus was a smaller ornithuromorph.
Classifying it has been tough as its fossil showed morphological similarities
with enantiornitheans and ornithuromorphs. However, further studies
conclusively put it in as a basal ornithuromorph as it was overall more derived
than earlier forms. Yet, coming from 130 mya strata does push ornithuromorphs
back another 5 million years plus.
Some ornithuromorph features in Ambiortus are: possessing a rectangular
medially projecting procoracoid, coracoid with a lateral process, U-shaped
furcula, curved scapula, rounded deltopectoral crest and a sternal keel just
reaching the rostral margin of the sternum. It was closely related
morphologically to Apsarvis.
The ornithuromorphan family, Hongshanornithidae (Hun-shawn-or-nith-ah-day)
had a temporal range of 130.7-120 mya. Hongshanornithids were small ranging in
size from 10.2-11cm/26-28in which includes the tail. The legs were long in
comparison to the body and most likely were shorebirds. All possessed a snout
with teeth mostly in the upper maxilla and lower dentary and still possessed
forelimb claws. As far as relations to other ornithuromorphs, hongshanornithids
were most closely related to the other ornithuromorph family, the songlingornithids.
There are five holotypic genera in the
family, Hongshanornithidae and they
are: Archaeornithura (Are-kay-ore-nith-ura)
from 130.7 mya; Tianyuornis (Tea-an-you-or-niss)
from 125 mya; Hongshanornis (Hun-shawn-or-niss)
for which the family is named after, from 122 mya; Longicrusavis (Lawn-guh-crew-say-viss) from 122 mya and Parahongshanornis (Pah-rah-hong-shawn-or-niss)
from 120 mya. All lived during the Early Cretaceous.
The fossil remains of Hongshanornis were collected in the
lacustrine deposits of the ‘Yixian Formation’ of Liaoning Province, China. With
a body and tail length of 18cm/7.1in and a wingspan of 32cm/12.6in, it was on
the smaller end in ornithuromorph size. The fossils are detailed in showing miniscule
teeth impressions in the front of the mouth and one small thumb claw on each
wing. Some of the fossils still retain bone tissue and feathering. Even though
the tiny rounded teeth and claws show a persistent remnant of ornithuromorph
dinosaurian ancestry, the reduction in amount and size prove that ancestral
theropodal dinosaurs had radiated out in setting the anatomical plan of birds.
 |
| Artist: Jack Wood Hongshanornis |
 |
| Hongshanornis fossil |
Hongshanornis most likely had
a generalist diet relying on insects, invertebrates along shores of aquatic
environments, invertebrates living within the shore mud and with the evidence
of gastroliths in one fossil, hints to the fact that it supplemented its diet
with plants.
 |
| Artist: Stephanie Abramowitz Longicruavis |
 |
| Longicrusavis Fossil |
Songlingornithidae (Song-lin-gor-nith-ah-day)
family members had a temporal range of 125-75 mya. Songlingornithids are
characterized by having a pair of medial fenestrae (openings) in the caudal
margin of the sternum. The metatarsals are also fused into a tarsometatarsus (fused
lower leg bone of birds and some non-avian dinosaurs). Also, songlingornithids
possess on the premaxilla at the tips of the lower jaws ossicles (tiny bones)
of unknown functional significance. Most all songlingornithids come from the ‘Jiufotang
Formation’ of Liaoning Province, China with one Yanornis species coming from the ‘Yixian Formation’ and Hollanda coming from Mongolia’s ‘Barun
Goyot Formation’. Only Hollanda comes
from the Late Cretaceous, the rest come from the Early Cretaceous.
There are four songlingornithids and
they are: Yanornis (Ya-nor-niss) from
124-120 mya; Songlingornis
(Song-lin-gor-niss) from120 mya; Yixianornis
(Yix-e-an-or-niss) from 120 mya and Hollanda
(Ho-lawn-duh) from 75 mya.
 |
| Artist: Jack Wood Yanornis |
 |
| Yanornis fossil |
 |
| Yanornis fossil with predigested fish |
 |
| Artist: Luis Chiappe Hollanda |
This songlingornithid appearing towards
the end of the Late Cretaceous shows that basal/primitive ornithuromorphs, even
though they all went extinct by the end of the Cretaceous, held a long
succession rate due to their evolutionary ability to fill ecological niches
whether it was in the air, water or ground. Through natural selection, they
adapted quickly in the geological record. Hollanda
was a ground dweller and likely spent most of its time near bodies of water in
a semi-arid environment in search of small vertebrates like lizards and
invertebrates like large insects. It would run before it would fly away from
danger.
Initially, Hollanda was considered more advanced and placed as a sister group
to Ornithurae, but later studies
proved it was phylogenetically related to the more primitive ornithuromorphs as
a basal songlingornithid.
Ornithurae is Greek
meaning ‘bird tail’ and includes the common ancestor to Ichthyornis, Hesperornis,
Limenavis, all other basal holotypic
ornithurans and of course all modern birds (Neornithes).
The ornithuran temporal range is from 121 mya to the Present. Most all
ornithurans were aquatic.
Ornithurans have autapomorphies that are
distinctive features as derived phylogenetic traits originating from the Ornithurae common ancestor. The ornithuran
cladogram includes all taxa more closely related to extant birds than to Archaeopteryx morphologically,
physiologically, phylogenetically and for sure, anatomically.
Most ornithurans had reduced wings that
could not support aerial transit opting to forego flight for an aquatic life.
The wings were used for paddling or held tightly up against the body for diving
to lessen resistance then utilized for maneuvering underwater. All ornithurans
either had lobed or webbed feet for aquatic propulsion in being the first line
of dinosaurs to enter into marine environments.
All the ornithurans’ adaptations to an
aquatic life indicate a long evolutionary history prior to their Late
Cretaceous life. As a monophyletic clade, ornithurans, even though they were
water fowl in both freshwater and marine, they are the closest relatives to all
living birds.
The Late Cretaceous toothed ornithuran
birds, Ichthyornis and Hesperornis fossils preserve remains
that are essentially indicative of modern bird bone tissue demonstrating the
fact that the unique growth strategy of living birds evolved during the
Cretaceous before Neornithes.
Furthermore, histological analysis of most basal birds from the Early
Cretaceous (ex: Sapeornis, Jeholornis) and basal ornithuromorphs from
the Late Cretaceous reveal slower forming bone tissue and lines of arrested
growth (LAGs) indicating that the fully modern growth strategy evolved within
the clade Ornithuromorpha’s early
basal representatives.

Enaliornis
(E-nal-e-or-niss) from 99.6-93.5 mya;
Pasquiaornis
(Pas-qwee-a-or-niss) from 93 mya with two species;
Baptornis (Bap-tor-niss)
from 83.5-80.5 mya;
Brodavis (Brod-a-vis)
from 80.5-66 mya consisting of four species;
Judinornis
(Ju-dee-nor-niss) from 70 mya;
Potamornis
(Poe-tah-mor-niss) from 66 mya;
Hesperornithidae (Hess-per-or-nith-ah-day)
from 85-82 mya with five genera ⇒
Fumicollis
(Fu-me-col-liss) lived between 86-82 mya;
Parahesperornis
(Par-ah-hess-per-or-niss) lived between 85-82 mya;
Hesperornis
(Hess-per-or-niss) from 82-78 mya;
Asiahesperornis
(A-Shah-hess-per-or-niss) from 70 mya:
Canadaga
(Can-ah-day-gah) from 67 mya.
Finally, in rounding out Ornithurae there is one holotype sister
group, Limenavis (Lye-men-nay-viss)
to Neornithes (Nee-or-nuh-theez) or Aves (Ah-vez), whichever name you
prefer.
 |
| Artist: Jack Wood Ichthyornis |
At 24cm/9.4in long and a wingspan of
43cm/17in, it was the ecological equivalent to present day seagulls and
skimmers. This most basal ornithuran is the only ornithuran to have flight
wings. With teeth only in the middle section then back towards the end of the
mouth of the upper and lower jaws, the toothless jaw tips functioned as a bird
beak. This allowed it to bite like a little dinosaur and peck like a bird. Teeth
shape was more flattened than rounded with a broadening base while the tips
lacking any serrations slightly curved backwards.


 |
| Ichthyornis skull & Skeleton |
 |
| Artist: Jaime Chirinos Ichthyornis stealing fish |
 |
| Artists: Mike Hanso, BA Bhullar Ichthyornis |
 |
| Artist: Masato Hattori Chupkaornis |
Hesperonithes was a highly
specialized clade of aquatic hesperornithiforms that inhabited marine and
freshwater environments. They were so specialized that the hind limbs developed
for propulsion through liquid mediums would have been useless for walking on
land and most likely were used only for shoving the body ventrally while on
terrestrial environments. Even the hip and knee joints were shaped such that hesperornitheans
could not move them dorsoventrally. The bones were dense making the animal less
buoyant for diving purposes. Morphologically, hesperornitheans had similarities
closest to modern day diving ducks and grebes.
The snouts were long and narrow, further
narrowing towards the tip which was tipped with a slightly hooked beak. Inside
the mouth the jaws were lined with small sharp teeth set in a longitudinal
groove unique only to hesperornitheans. The beak tip was toothless. All
hesperornitheans come from the Late Cretaceous and diverged from their and
birds common ancestor during the Early Cretaceous.
 |
| Artist: Nobu Tamura Enaliornis |
There is no evidence of a toothed snout
and the wings with a length from shoulder to the distal end of the body, shows
signs of the beginnings of atrophying in displaying vestigial evolvement toward
diving purposes as opposed to flying. The toes, although not webbed were
heavily lobed as the shapes of the distal tarsometatarsal trochlea (an
anatomical structure acting as a pulley) present.
 |
| Enaliornis Braincase air pockets |
 |
| Artist: Gunar Björkman Baptornis |

Baptornis existed during the rise in ocean waters creating North America’s large inland sea, the ‘Western Interior Seaway’ that stretched from the Gulf of Mexico through the Midwestern U.S. and Canada ending in the Arctic Ocean. Existing from the mid-Cretaceous to the earliest of the Paleocene, this interior seaway was home to Baptornis. With fossil remains found in Sweden where the former, ‘Turgay Strait’ (also known as the, ‘Turgai Sea’) once met the ancient North Sea, Baptornis also occurred there. In fact it may have swam and dove the entire Holarctic.
 |
| Baptornis skeletal Martin & Tate et al |
Baptornis consists of
numerous adults and juveniles. They have been found along ancient coastal
shorelines and deep out into ancient sea sediment. This suggests that this
basal hesperornithean was migratory or at least frequented northern islands to
breed as that is where most of the juvenile fossils come from.
 |
| Artist: Matthew Martyniuk Brodavis |
Although there is very little evidence
of pachyostosis (bone thickening) to negate buoyancy, anatomical features still
point to a diving existence along with lobed feet. With thinner bones than its
hesperornithean relatives and wing bones that had not yet begin to radically reduce
in size as nowhere near a vestigial stage, Brodavis
most likely was volant (could fly). All these examples add up to the fact that Brodavis could have lighter bones in
freshwater environments because freshwater is less buoyant than seawater and
with lakes as much smaller bodies of water than seas, it could also frequent
other lakes through flight to satisfy its piscivorous diet.
All members in the family, Hesperornthidae
evolved for a specialized aquatic life along coastal sea shores as well as into
deeper offshores. The hind limbs were adapted to propel through aquatic
environments, while the wings had become vestigial being incapable of flight.
On land however, the hind limbs are attached to the sides of the ilium and
would be useless for walking, so at best hesperornithids scooted across solid
terrain on their bellies.
Most hesperornithid species come from N.
America, but there have also been recent discoveries in Sweden and Kazakhstan
where the ‘Turgay Strait’ once was and a very recent 2018 Saratov Province,
Russia find of H. rossicus from the sandstone of the ‘Campanian
Rybushka Formation’.
 |
| The Turgay Strait |
 |
| Artist: Jaime Chirinos Hesperornis |
Hesperornis species were
rather large averaging ~ 1.8m/5.9ft in length. Although they possessed a
keratinized beak, except for the premaxilla being edentulous (toothless) the
mouth was rowed with small sharp conical teeth. The beak was used in snatching,
while the teeth were utilized in grasping slippery marine prey. The beak had
small pits in the upper palate that allowed teeth from the lower jaws to lock
into the pits preventing the beak from opening disavowing any thrashing prey to
escape.
 |
| Artist: John Conway H. regalis |
 |
| Artists: Julio Lacerda & Henry Sharpe Hesperornis as thief & prey |
Hesperornis was a true
piscivore predator, but no doubt was an opportunist in scavenging washed up
dead fish and even torment other animals in stealing their fish prey. It appears at times however that Hesperornis was itself on the menu. Discovered
in the ‘Pierre Shale Formation’ of South Dakota, among other predigested
animals, the partially digested remains of Hesperonis
was found in the gut cavity of a large Tylosaurus
proriger fossil.

 |
| Hesperornis fossils |
The Hesperornis
genus countered its avialan ancestry that was trending in smaller sizes to
accommodate flight. In foregoing flight for an aquatic lifestyle, later
hesperornithids began increasing body size and one of the last hesperornithid
species to appear was Canadaga was
the largest one of all in the lineage to modern birds. From 1.5m/4.9ft to a
whopping 2.5m/8.2ft, Canadaga played
second fiddle in size to no other. Almost half that size was the neck.
 |
| Credit: Canadian Museum of Nature Canadaga |
 |
| Canadaga vertebrae A)Ventral B) Dorsal |
Before venturing into Neornithes, the last ornithuran we’ll
discuss is Limenavis (Li-men-nay-vis)
in which the 70 mya fossil comes from the ‘Allen Formation’ at Salitral Moreno,
Argentina. The fossil has been difficult to classify. The remains, definitely of
a bird, were originally put down as a basal ornithuran. However, later analyses
found it to be much more advanced than Ichthyornis
and classified it as a carinate bird (birds possessing a keel or ‘carina’).
Thus, as one of the first true birds, scientists gave it the name, Limenavis in meaning, ‘threshold bird’.
Based on the results of the phylogenetic
analysis and identification of autapomorphies, Limenavis is now considered to be the closest species to the common
ancestor of the crown clade, Neornithes
of all living birds. Some paleontologists are even suggesting its anatomical
features may put it as a direct common ancestor linked to paleognathes.
 |
| Artist: Alexander Lovegrove Limenavis being flushed |
Neornithes/Aves:

Neornitheans are the most recent common
ancestors to modern birds including all their descendants extinct and extant. Neornithes is divided into the two
superorders: Palaeognathae (Pay-lee-og-nuh-thee)
consists of the flightless ratites and the tinamous and Neonagthae (Nee-og-nuh-thee) consists of all other extinct and
extant flying birds. The ratites include birds like the ostrich, emu,
cassowary, rheas, kiwi, the extinct moa, elephant bird and the extant family, Tinamidae (Tin-ah-me-dee). Tinamous are
capable of flight (albeit poor fliers), but are more closely related to
ratites, especially in possessing the more primitive reptilian palate in the
upper mouth as the ratites have.

 |
| Neornithean Cladogram Top: Paleognaths Bttm: Neognaths |
 |
| Artist: Franco Tempesta Chicxulub Imoact |
 |
| The Cenozoic Era |
No tetrapod weighing over 25kg/55lbs survived beyond the extinction boundary. With bird lineages decreasing in length, this smallness is what most likely aided birds in their survival beyond the extinction boundary. Able to hide in small burrows, tree holes or shelter in what marsh and wetland environments offered, gave small neornitheans a concealed advantage to extreme exposure limits. Within 10,000 years, neornithean survivors soon after the extinction, dispersed and radiated out filling ecological niches left vacant by those species that went extinct.
Along with birds, 50% of crocodyliforms 80%
of Cretaceous turtles, the order Squamata
(lizards and snakes), freshwater tetrapods such as amphibians and all the small
mammalian lineages except the Asian forms survived through the K-Pg extinction
event.
Palaeognathae: Palaeognaths,
meaning ‘old jaws’, as just mentioned above is due to the fact that they share
a primitive bony palate (mouth roof). Their temporal range is from ~ 100mya at
the beginning of the Late Cretaceous to the Holocene of the present. The palate
is defined with five unique distinctions and they are:
1)
The
large vomer articulates with the premaxillae and the maxillopalatines
anteriorly, while the vomer fuses to the ventral surface of the pterygoid with the
palatines fusing to the ventral surface of this wholly pterygovomer
articulation.
2)
The
palatine is also fused into a rigid joint with the pterygoid.
3)
This
fusion with the pterygoid prevents the palatine from articulating medially with
the basisphenoid.
4)
Articulation
on the pterygoid for the basipterygoid process of the basicranium is diverted
to the articulation between the pterygoid and quadrate.
5)
The
pterygoid/quadrate articulation includes the quadrate’s orbital process.
Palaeognaths also share a similar pelvis
anatomy and patterned grooves in the keratinous covering of the beak. For most
palaeognaths, the male is the one who incubates the eggs laid down by the
female in the nest he arranged. More than one female may lay eggs only in one
male’s nest and she may have been fertilized by another male. Only in ostrich
species and one species in the genus Apteryx
(App-ter-rix), the great spotted kiwi does the female assist the male in
incubation. Most palaeognaths have lost their keel.
Except for kiwis’ exceptional brain/body
ratios, paleognathes have very small brains compared to body size. Palaeognaths
are known for their large sizes with the extant ostrich reaching 2.7m/9ft in
length and emus up to 2.3m/7.5ft. But kiwis’ range of sizes is only
36-56cm/14-22in, while the dwarf tinamou is only 15cm/6in in total length. When
it comes to mobility, the larger palaeognaths use speed with ostriches capable
of reaching 60kph/35mph while the extinct bush moa, with a walking speed of 3-5kph/1.75–3mph,
could sprint up to 69kph/43.9mph.
There have as yet been no Cretaceous palaeognath
fossil finds, but we do know that the extant five species of kiwis (Apteryx of New Zealand), the three
species of cassowaries (Casuarius of
northeastern Australia and New Guinea), the emu (Dromaius of Australia), the two species of rheas (Pterocnemia and Rhea of South America) and the ostrich (Struthio of Africa) are all closely related.
 |
| Continental Drift |
 |
| Gondwana breakup Griem 2007 |
However, there is also a controversial
analysis that has become the prevailing thought due to its undeniable bare
knuckle molecular genetic results. Due to recovered nuclear genome fragments
from extinct elephant birds, identified homoplasies (singular: homoplasy ~ independently
a gained or lost trait) in morphological traits of palaeognaths and reconstructed
morphology based phylogenies, including fossil species evaluations provided
stable divergence time estimates enabling a valid argument regarding
correlation with geological events.
So, a few researchers have proposed a
new evolutionary scenario from this data that contradicts the traditional view.
The ancestral Palaeognathae were
volant (capable of flight), as estimated from their molecular evolutionary
rates and originated during the Late Cretaceous first in the Northern
Hemisphere. They migrated aerially to the Southern Hemisphere and explosively
speciated around the K-Pg boundary. They then extended their distribution to
the Gondwana derived landmasses, such as New Zealand and Madagascar by overseas
dispersal. Gigantism subsequently occurred independently on each isolated
landmass.
The earliest palaeognath species thus discovered come from the Paleocene and Eocene 61.7-0 mya and are classified under the two families, Lithornithidae (Lith-orn-ah-day) from 61.7-40 mya and Rheidae (Ree-ah-dee) from 56-0 mya, in which rheids are still around today in South America. Also, from ~ 58.7-48.6 mya, Diogenornis (Dye-oge-en-orn-nis) was a stem group casuariiform related to extant cassowaries and emus as well as a sister group to lithornithids.
The earliest palaeognath species thus discovered come from the Paleocene and Eocene 61.7-0 mya and are classified under the two families, Lithornithidae (Lith-orn-ah-day) from 61.7-40 mya and Rheidae (Ree-ah-dee) from 56-0 mya, in which rheids are still around today in South America. Also, from ~ 58.7-48.6 mya, Diogenornis (Dye-oge-en-orn-nis) was a stem group casuariiform related to extant cassowaries and emus as well as a sister group to lithornithids.
Lithornithid fossils come from the Upper
Paleocene to the Middle Eocene in what is now North America and Europe. They
all were volant (capable of flight), although with long hind limbs and strong
tridactyl feet, some were more prone to running before resorting to flight.
Lithornithids are distinctive palaeognaths in retaining a fully developed
hallux (innermost digit of hind foot, or big toe) unlike their palaeognath descendants
whose feet evolved for running speed. Most lithornithids possessed perching
feet, well developed keels, pectoral girdle elements and proportionally large
wings allowing for soaring flight akin to that of modern birds such as storks.
Once the Pleistocene arrived heralding
in the ‘Ice Age’, all the subtropical/tropical climates and environments left
the North American continent and with that period change, so too, lithornithids
followed suit in becoming extinct.
There are five genera of lithornithids
and they are: Fissuravis (Fis-sure-a-vis)
from Germany’s ‘Walbeck’ fissure strata 61.7-58.7 mya; Lithornis (Lith-orn-nis) from Montana’s ‘Bangtail Quarry’ 56-40
mya; Paracathartes (Pair-ah-cath-art-teas)
from Wyoming’s ‘Willwood Formation’ ~ 55mya; Pseudocrypturus (Sue-doe-crypt-ur-us) from Wyoming’s Green River
Formation’s ‘Fossil Butte Member’ 52mya and Calciavis
(Cal-see-a-vis) from Wyoming’s ‘Green River Formation’ 51 mya. The ‘Green River
Formation’ of Wyoming is composed of lake bed sediment and is the richest
former lake bed for fossils worldwide with its varve layers presenting a
continuous six million years record.
 |
| Artist: Velizar Simeonovski Calciavis |
 |
| Artist: Piotr Gryz Pseudocrypturus |
 |
| Artist: Scott Reid Paraacathartes |
Paracathartes was different
among the other lithornithids in having shorter and more robust wing elements
and a deeper keel relating to a flight style more akin to that of extant
tinamous making it far more terrestrial than the other members. With this in
mind, Paracathartes could be the
intermediate between the other lithornithids and the flightless ratites of
today, or it simply is an isolated example of convergent evolution.
 |
| Artist: Geraldo Diogenornis |
As in the extant cassowaries and emus, Diogenornis’ maxilla ended in a point, the tibiotarsus (bird’s leg bone corresponding to the tibia that’s fused at the lower end with some tarsus bones) with the condyles medalis (a rounded articulating protuberance on the inner side of the lower extremity of the femur) aren’t undercut proximally and the shape of the trochlea fibularis (a projection from the lateral side of the calcaneus between the tendons of the peroneus longus and brevis) was large and oval. Along with these shared traits coming from Paleocene South America, Diogenornis’ relations to Australian ratites corroborates a Gondwanan origin of these birds and the close relationship among the Causauriiformes in the extant Rheidae (rheas), Casuariidae (cassowaries), and Dromaiidae (emus) families along with the importance of the Transantarctic biotic interchange of birds between South America and Australia in the beginning of the Cenozoic.
 |
| Credit: Izaat Design Lmtd. Moa |
Moas, as ratites were flightless, but in
addition did not even have vestigial wing remnants as all other ratites did and
do. In reproduction, moas laid one 18cm/7in sized egg into a scratched out
earthen nest. Laying just one egg and in the addition of studies on the moa
cortical bone growth rings, alludes to the fact that moas were K-selected.
Species strategies in r/K selection are the concerned modes of successful
reproduction utilizing combinations of traits that trade-off between quantity
and quality of offspring. In having many offspring with minimal parental care
is r-selection. K-selection is in having minimal offspring with much more attended
parental care.
 |
| Artist: Sameer Prehistorica Man compared to Moa |
 |
| Artist: John Megahan Haast’s eagle attacking a pair of moas |
 |
| Moa foot |
 |
| Palaeognaths: kiwi, ostrich, moa skeletons |
The elephant bird of Madagascar was
another huge palaeognath that also went extinct due to man once he arrived on
the island. There were three genera in, Aepyornis
(Eep-pee-or-nis) Mullerornis
(Mule-lur-or-nis) and Vorombe (Vo-rom-be),
Perhaps the largest, but most definitely the heaviest bird ever, Vorombe stood 3m/9.9ft tall and weighed
730kg/1,600lbs.
 |
| Artist: Brian Choo Elephant Bird (Aepyornis) |
According to the mtDNA (mitochondrial DNA) analyses and hybridized enrichment of
in-solution RNA arrays of elephant birds, they did not evolve as isolated from
an original population during the breakup of Gondwana, but rather via the
flight migration of a direct ancestor originating from New Zealand some 50 mya
ago well after New Zealand had rifted then drifted from Australia.
 |
| Vicariance vs. mtDNA in distribution |
Molecular dating uses differences in DNA sequence from one species to another to estimate how long ago the species diverged. This type of technique accounts for genetic changes or mutations that accumulate over a rated time. Thus, the greater the difference in genetic sequence, the farther back the divergence of the speciation dates.
In the illustration below, also from Dr.
Mitchell’s team, shows species divergence and phylogenetic position of the
elephant bird from the mtDNA
sequencing. In mya the blue numbers are the time of divergence. Blue arrows
mark the minimum date for the evolution of a flightless lineage as evidenced in
fossils and the species description in red are those that are extinct.
 |
| Palaeognath genetic positioning |
 |
| Elephant bird egg compared to a chicken egg |
 |
| Elephant bird egg compared to an adult kiwi |
 |
| Cast of elephant bird wingbone |
So, in setting up a biogeographical
layout, these charred egg fragments show that the eggs were transported then
cooked over a fire. The commonality of these fragment study findings indicates
that these early people relied heavily on the eggs, but in coexisting for
thousands of years, did not hunt adults that extensively. Perhaps due to a
cyclic climate change or a depletion of natural habitat, but not this time
solely due to predation by man did the elephant bird go extinct with the last
one expiring by the mid-17th century.
Current palaeognaths, as ostriches and
rheas have long wing feathers that attach to long tridactyl two-clawed hands.
Emus and cassowaries have very short wings with a substantially reduced
single-clawed carpometacarpus and poorly ossified (fused) splint for the alula.
Kiwis have stick-thin wings where the manus (hands) are monodactylous (one
digit), elephant birds are the same as kiwis but thicker and of course much
larger, while moas had no wings at all.
Of the extant palaeognaths, rheas are
the basal most branch. Tinamous and moas share a common ancestor, with kiwis
more closely related to emus and cassowaries. Also as previously stated, pretty
much the largest bird ever known, the extinct elephant bird of Madagascar has
for its closest relative, the smallest extant palaeognath in the kiwi. The
sister group to all modern palaeognaths is the ostrich family, Struthionidae (Strut-the-on-ee-dye). The
overall direct line to a common ancestor for palaeognaths may go back to 120
mya.
With two extant species, rheas come from
South America and have a temporal range of 126 thousand years ago to the
present. However, in the family, Rheidae
(Ree-ah-dye) there are extinct species and subspecies that go back 56 mya, such
as R. fossilis and R. pampeana. Though somewhat smaller, rheas
are very similar in appearance to ostriches, their closest relations.
As a ratite, they are flightless, but
use their reduced wings for balance and act as an airfoil for changing
direction. The legs are used for running speed in utilizing the legs length and
muscle strength. Rheas are omnivorous in dieting on primarily broad-leafed
plants, fruit, seeds and roots while complementing the diet with insects like
grasshoppers and carrion.
Males are polygamous, mating with two to
twelve females and after courtship, builds a nest by scratching a pit in the
ground layering it with grass and leaves. The females then lay up to 60 of their
fertilized eggs in the nest where the male incubates and tends to the
hatchlings. In fending the hatchlings, males will charge at anything he deems
threatening to his brood.

There are nine extant genera of tinamou and have a temporal range of 10 mya to the present in quite possibly having lithornithids as being directly ancestral to. The earliest decisive tinamiform fossil found was in Argentinean Patagonia during the Miocene’s Burdigalian Stage 17.5-16.3 mya of the genus, Crypturellus (Krip-tu-rail-us). The extant species come from Mexico, Central and South America.
Like most extant palaeognaths, tinamous
are ground birds and as previously stated the male tends to the eggs and
hatchlings. But the females lay some unique, fancy and pretty eggs into the
male’s nest. Unlike most bird eggs, tinamou eggs are glossy and show hues of
vivid color. The reason being is that they are smooth instead of pitted and do
not get color from pigmentation, but from iridescence.

Along with the eggshell being made up of calcium carbonate, the last thing the female does prior to laying her egg is coating it with a layer of calcium phosphate giving the egg its smoothness and iridescence. The smoothness gives the glossy appearance and the iridescence gives it the colorful hues. At differing angles, the normal blue color underneath can turn shades of varying colors. No one knows the underlining reason for this tinamou egg attribute, but the ones that did the egg testing and analysis feel that it may aid in attracting females to lay their eggs in a nest with eggs giving off their colorful hues laid previously. With more eggs in a nest, it might be a strategy that more will succeed in hatching due to a lessening of predation if very few eggs were only in a nest.
Neognathae: Neognaths
possess a differing jaw structure than paleognaths, so the term refers to ‘new
jaws’ even though the palaeognath jaw is apomorphic (more derived) between the
two and is not indicative to comparative evolution. With over 10,000 extant
species, Neognathae covers virtually
all living birds from hummingbirds and penguins to vultures and eagles along
with their direct common extinct ancestors.
 |
| Credit: marysrosaries Neognath upper jaw vs. Palaeognath |
1)
Loss
of the basipterygoid articulation with the cranium.
2)
A
developed process of the pterygoid/palatine joint.
3)
Reduced
vomer, or lost entirely doesn’t reach caudally to the pterygoid.
Molecular dating points to a neornithean
Cretaceous radiation where the lack of Cretaceous fossils suggests a Paleocene
radiation after the K-Pg extinction. Except for a few near-complete articulated
fossils, but mostly a spattering of some isolated bones, there has been some
concern sprinkled with a dose of consternation as to why there hasn’t been a
trove of Neornithes Cretaceous fossil
finds. Neornitheans should’ve been well established at least by 95 mya leading
into neognaths.
As has been suggested by paleontolgists,
Roger Cooper and Richard A. Fortey, some neorinthean clades did originate deep
in the Late Cretaceous but maintained a low Mesozoic diversity. Also, paleo-ornithologist,
John L. Cracraft states that the lack of neornithean fossils before the K-Pg
extinction event could be the case that much of Cretaceous neornithean
evolution took place in the Southern Hemisphere, where the fossil record has
been far less thoroughly sampled. For whatever is the reason, I’m quite sure of
it that in the not so distant future with more fossil finds and sharp minds,
this conundrum will be solved. Anyway, there is fossil evidence of an explosive
diversification of modern bird families to fill in the biological niches left
vacant after the K-Pg extinction.
 |
| Artist: Monica Serrano Vegavis being chased |
 |
| Artist: Nicole Fuller Sayo Vegavis syrinx |
The syrinx has an asymmetrical third
segment to it and with the soft tissue acting as a resonator, suggests that Vegavis was capable of producing deeper
toned honks just as ducks and geese do today.
 |
| Vegavis scanned syrinx |
Birds can also harmoniously sing without
interruption in taking a breath, due to their one way entry and exit air
passages where there is always a constant flow of fresh oxygenated air. In
humans, breathing is much more imperfect. When we exhale and begin to inhale,
we actually inhale a bit of the spent exhaled air, contaminating the fresh
inflow of oxygenated air. For this and other reasons as dealing with trachea
musculature, the human larynx converts only 3% of energy spent into sound where
the bird syrinx, as far more efficient, converts 100%.

The advancement witnesses in the Vegavis fossils attest that neognaths were well on their way to modern bird forms before the K-Pg extinction, implying with physical fossilized proof that at least some groups of neognath modern day birds first came from the Cretaceous.
Genyornis
(Gee-nye-or-niss) also belonged to the duck/goose clade of anserimorphs, which
mainly represents extinct and extant water fowl. However, at 2m/6.6ft in length
and weighing ~230kg/507.1lbs, this was one big goose on steroids. With a
temporal range of ~ 126-30±5 kya (thousand years ago) in what is now Australia
appears to have simply vanished after 30,000 years.
With vestigial wings, Genyornis was flightless relying on its
strong leg muscles and stout hind limbs for rapid mobility. Unrelated to other large
flightless birds, Genyornis had a
short dentary symphysis and a dorsally directed pterygoid process on the
quadrate, a squarish bone that is articulated by the jaws and homologous with
the incus bone of the middle ear in mammals. Also, eggshell studies show
similarities to megapodes (Australasian land fowl, such as mallee fowl).
With fossils and eggs found in
collective sites, Genyornis was
social. Studies on Genyornis eggshell
fragment piles show that they were heated or cooked on one side going as far
back as 47,000 years ago and too, there are ancient aboriginal stone art drawings
of Genyornis preserved still today.
 |
| Artist: Peter Trusler Genyornis & Megalania |
Palaeognath ratites weren’t the only large
flightless birds, for as in Genyornis,
neognaths also had their fair share of big ‘uns…and no, ‘Big Bird’ of Sesame
Street fame doesn’t count. Once the reign of dinosaurs were over, other
reptiles, mammals and the last of the dinosaurs in birds were now able to fill
the void in ecological niches in which for some, gave latitude for size
increases. Below are a few more huge
neognaths.
 |
| Artist: Jaime Chirinos Dromornis |
The temporal range for Dromornis is 8 mya-30 kya, but the Australian
family, Dromornithidae
(Dro-mor-nith-ah day) had a temporal range of 25 mya-3 kya. Dromornithids are
in the genera, Genyornis, Dromornis, Barawertornis (Bah-rahweir-torn-iss), Bullockornis (Bull-loc-corn-iss) and Ilbandornis (Ill-ban-dorn-iss). The earlier dromornithids were much
smaller than the latter as in, Barawertornis
having a total length of ~ 1.7m/5.6ft.
In Dromornis,
the species was dimorphic with the males more robust, stout and heavier than
the females, but both sexes were much the same size in length. With the huge
and powerful beak, some earlier research statements inferred that Dromornis was a carnivore, however later
analyses proved the beak lacked carnivory features while showing features
favorable to an herbivory diet. This bird was indeed a generalist herbivore
eating fruit, nuts, various plant foliage and roots.
 |
| Artist: Taringa Gastornis |
 |
| Gastornis skeletal |
Another snag in Gastornis is that again, since it was big with a huge powerful
beak, initially it was thought it must have been carnivorous going around the
forests chasing down little mammals and gobbling them up. But, a carnivory
biochemical jaw analysis did not pan out as so and Gastornis possessed an herbivore foot with no carnivore claws,
while calcium isotope studies of its bones show no evidence of it in being a
meat eater.
The eggs of Gastornis were unique in that instead of being round or even oval,
they were oblong and measured 25.4cm/10in long by 10.2cm/4in in diameter.
Besides the numerous anatomical fossil remains, also discovered have been its
feathers from a ‘Green River Formation’ fossil find and footprints from France
and Washington State, USA.
 |
| Credit: dinoanimals.com Phorusrhacos |
Phorusrhacos, with greatly
reduced wings was flightless and stood on long stilted-like legs looking
somewhat like an extant red-legged seriema with a huge beak. Its vestigial
wings had the forelimbs manus ending in a hooked claw. Ya might note that the
generic name doesn’t end in ‘ornis’, which is Greek for ‘bird’, as most other
large flightless neognaths do. That is because the first Phorusrhacos fossil discovery in 1887 consisted only of a partial
lower jawbone and was mistakenly identified as coming from a mammal. That was
soon corrected in 1891 after more fossil finds collaborated in what the jawbone
really came from, but the generic naming stuck.
 |
| Phorusrhacos skull |
But with mid-size to smaller prey, I
would like to include that Phorusrhacos, with its long legs may have pounced onto the prey and while bearing down, with
its weight sinking claws into flesh, pinned the prey securely to give it a few
shots with the pointed tipped beak. This is how the extant secretary bird
catches its snake prey and with the long legs in both birds, it keeps the body
away from the dangers of a struggling or defensive prey. Phorusrhacos most likely would not have passed up the opportunity
to scavenge any carrion it came by either.
Nonetheless, the wings’ hooked claws
would not have been too much of an advantage in seizing prey as it would’ve
brought the head and torso too close to a defensive prey’s retaliation. The
wings’ hooked claws if used at all would’ve been for combating a fellow
competitor during mating or territorial rights.
 |
| Artist: Jaime Chirinos Titanis |
In fact there followed many other fossil
finds that closely resembled Phorusrhacos,
but with a few distinguishing features it soon was revealed the super family, Phorusrhacoidea
(Foe-roos-ray-koi-dee-ah) was needed and devised. With 18 genera of ‘terror
birds’, below is the member listing under the superfamily, Phorusrhacoidea:
 |
| Credit: Manasataramgini Terror Birds |
Superfamily: Phorusrhacoidea
Genus: Lavocatavis – Middle Eocene Glib Zegdou
Formation of Algeria
Family: Phorusrhacidae
Genus: Patagorhacos – Early Miocene Chichinales
Formation of Rio Negro Province, Argentina.
Subfamily: Brontornithinae — species standing over
2.3m/7.5ft high. Some monophyly placement dispute in Phorusrhacidae
Genus: Brontornis [Early to Middle Miocene
(Santacrucian-Laventan) Santa Cruz and Monte León Formations, Argentina]
Genus: Paraphysornis [Late Oligocene to Early
Miocene (Deseadan) Tremembé Formation of São Paulo State, Brazil]
Genus: Physornis [Middle to Late Oligocene
(Deseadan) Sarmiento Formation of Santa Cruz Province, Argentina]
Subfamily: Phorusrhacinae — species 3.3m/9.9ft high, but somewhat
slender and decidedly nimble
Genus: Devincenzia [Late Oligocene to Early
Miocene (Deseadan) Fray Bentos Formation of Uruguay]
Genus: Kelenken [Middle Miocene (Colloncuran)
Collón Cura Formation of Río Negro Province, Argentina; largest known
phorusrhacid]
Genus: Phorusrhacos [Early to Middle Miocene
(Santacrucian) Santa Cruz Formation of Argentina)
Genus: Titanis [Early Pliocene to Early
Pleistocene (Blancan) of Florida/Texas]
Subfamily: Patagornithinae — intermediate sized/very
nimble species, standing ~1.7m/5.6ft
Genus: Patagornis [Early to Middle Miocene
(Santacrucian-Laventan) Santa Cruz Formation of Santa Cruz Province, Argentina]
– includes Morenomerceraria, Palaeociconia, Tolmodus
Genus: Andrewsornis [Middle to Late Oligocene
(Deseadan) Agua de la Piedra Formation of southern Argentina]
Genus: Andalgalornis [Late Miocene to Early
Pliocene (Huayquerian) Ituzaingó Formation of northwestern Argentina]
Subfamily: Psilopterinae — species standing 70–100cm/2.3–3.3ft
Genus: Eleutherornis [Middle Eocene (Bartonian)
of Rhône, France and Baselland, Switzerland]
Genus: Paleopsilopterus [Middle Paleocene
(Itaboraian) Itaboraí Formation of Itaboraí, Brazil; identity as a phorusrhacid
dubious]
Genus: Procariama [Late Miocene to Early
Pliocene (Huayquerian-Montehermosan) Cerro Azul and Andalhuala Formations of
Catamarca Province, Argentina]
Genus: Psilopterus [Middle Oligocene (Deseadan)
Santa Cruz Formation and Late Miocene (Chasicoan) Arroyo Chasicó Formation of
southern and eastern Argentina]
Subfamily: Mesembriornithinae — medium-sized
species standing 1.2-1.5m/ 3.9-4.9ft
Genus: Mesembriornis [Late Miocene to Late
Pliocene (Montehermosan) Monte Hermoso Formation of Argentina]
Genus: Llallawavis [Late Pliocene
(Chapadmalalan) Playa Los Lobos Allo Formation of northeastern Argentina]
Below is a documentary on South American
‘terror birds’. At nearly fifty minutes it is a bit lengthy, but if ya have
time, it’s well worth it:
With a temporal range of 58.7-2.5 mya, the family, Pelagornithidae (Pal-ah-gor-nith-ah-day) consisted of seabirds dispersed all over the Late Paleocene-Early Pleistocene world. Fossils have been found in coastal rock from Belgium through Togo to Antarctica. Also known as false-toothed birds, pelagornithids had pointed tooth-like projections jutting from the edges of the beaks. Unlike true teeth and more like bone, these beak teeth contained Volkmann’s canals that are micro channeled tubular arrangements interconnecting haversian canals to one another. These tooth-like points were outgrowths of the premaxillary and mandibular bones.
 |
| Artist: Mark Witton P. chilensis |
 |
| Artist: Jose Vega a pelagornithid |
 |
| P. miocaenus skeletal |

As mentioned earlier above, P. sandersi belonged to the family of birds with the longest wingspans ever, while at, 6.1-7.4m/20-24ft it had the largest wingspan of any pelagornithid yet discovered. As in all pelagornithids, its false-teeth weren’t composed of enamel or any other hardened material, so were fragile as numerous fossils attest to in broken pseudo teeth. This alludes to the fact that food had to be soft-bodied like cephalopods or scaleless small fish. This would make good in utilizing the pseudo teeth for snatching prey just below the surface and swallowing whole, but not for tearing or chewing prey. As in being related to pelicans, a pelagornithid’s mouth was mechanized to open wide. Inside the eye sockets of some pelagornithid fossils, salt glands were evident, in particular in younger ones.
P. sandersi lived during the Oligocene 25
mya off the coast of what is now South Carolina, USA. It had short stumpy legs
ending in webbed feet with no hallux (big toe) and most likely hopped towards a
cliff whenever it was ready to take-off and become airborne. In performing
dynamic soaring, it has been estimated that it could reach speeds up to
70kph/37mph and filled the oceanic niche that pteranodons had left vacant.
In the direction of rooted phylogenetics,
basal penguins first show up as the most recent common ancestor 70-68 mya, a
few million years before the K-Pg extinction. This occurred within the regions
of what is now southern New Zealand and Byrd Land, Antarctica. Penguins didn’t
come directly from flying birds, but their basal ancestral flightless seabirds
did.
 |
| Credit: Hedwig Reunes-Vanhaevre |
Since they were now an oceanic bird,
basal penguins had begun the process of devolving back to heavier bones filled
with marrow. Thinner bones with air filled cavities for flight advantages were
no longer viable for a watery environment, but added weight for submergence and
countering buoyancy was.
The earliest penguin fossil finds combines
primitive and derived characters.
Incorporating fossil calibration points, DNA sequences, maximum likelihood,
and Bayesian analysis, penguin calibrations imply a radiation of modern (crown
group) birds in the Late Cretaceous 74 ± 3 mya.
 |
| Some prehistoric penguins |
 |
| Credit: Slack et al Waimanu tuatahi |
Icadyptes (I-kaah-diptz)
occurring in what is now Peru from the coastal desert marine rocks of the,
‘Otuma
Formation’ lived ~ 36 mya and was around 1.5m/4.9ft in length. It is the ancestral
apex sphenisciform in beak length possessing one that reached 20cm/8in long.
Still possessing the basal ancestral long pointed beak, Icadyptes’ beak was exceptionally long and is known as the spear-like
beak penguin.
 |
| Icadyptes beak comparison |
Palaeeudyptinae
(Pay-lee-oi-dip-sha-nee), fossils found from the Middle/Late Eocene to the
Middle Miocene is an extinct penguin sub-family group that had some of the
largest penguin members to ever swim the oceans. The fossils were discovered in
what is now: Antarctica, Australia, New Zealand and the western side of South
America. The decline and eventual extinction of Palaeeudyptinae species coincides with the competing forces of
cetacean and pinniped mammal groups and in addition, the coming of colder
climates and ice.
As a mid-evolutionary lineage Palaeeudyptinae species were more
primitive than today’s penguins in transitioning modern characteristics but
still retaining anatomical features from their seabird ancestors. While
ancestral wing feathering had already been lost, the semi-rigid flipper
assemblage had not occurred as in today’s penguins. For propelling capacity
while diving, the ulna (long bone in forearm) and radius (shorter bone in
forearm) had already flattened for increased capacity. The elbow and wrist
joints however, still retained degrees of flexibility as opposed to the rigidly
locked joint mechanisms in extant penguins.
 |
| Artist: Discott |

Where Anthhropornis was close to being as long as a VW Beetle automobile, Pachydyptes (Pac-he-dip-tees) was about as wide as one. This penguin was massive in girth with a weight of up to 100kg/220.5lbs, but a height of only 1.5m/5ft. This is not conclusive as the mass was calculated by only a few bones contained in the fossil find, but experts in reconstructing fossils agree that the estimate is close if not accurate. It’s not just from the bone sizes in determining the weight, but also in the wing bones, the width shows signs of being extremely stocky in girth. The wing bones are 35% longer than and nearly twice as thick as an emperor penguin’s.
We’ve gone over a lot of giant penguins,
but in the tiny Eretiscus fossil from
22 mya of Patagonia, Argentina, there was room for some small penguin niches as
well. Originally named Microdytes in
meaning, ‘tiny diver’ but named changed after it was found that a water beetle
had already been given that genus name. The current and now permanent name
change is, Eretiscus in meaning, ‘tiny
rower’. We’re only talking about millimeters here, or one-fortieth in inches
difference, but this penguin is around the size of the extant blue penguin at
46cm/18.1in and 1.5kg/3.3lbs in weight so, it may or may not be the smallest
penguin ever known, but it is currently the smallest penguin fossil ever known.
 |
| The Blue Penguin |
To answer the question, “Do penguins
have knees?”…yes they do; in fact the structural penguin leg configuration is
the same as that of a human leg consisting of a femur, tibia and fibula.
However, the femur is short and points outwards and upwards before joining the
tibia and fibula at the knee joint. With this configuration, it’s like a human
walking on stilts. Also, with the knee joint up so high, there is an illusion
developed by fur covering and hiding the joint and most of the leg making the legs
look very short and the gait appear as stiff legged.
 |
| The penguin knee |
 |
| The 61 mya penguin foot versus extant emperor penguin |
 |
| A hoatzin pair |
The oldest known fossils come from
Europe near Romainville, France. The fossil was found in sediment that was
deposited at the very end of the Eocene 34 mya. The genus name is Protoazin (Pro-toe-ah-seen). Namibiavis (Nuh-mib-e-a-viss) comes from
what is now Namibia, Africa and was found in Middle Miocene deposits laid down
16 mya. Genetic research alludes to the fact that the hoatzin is the last
surviving member of a bird line that branched off in a separate direction from
the rest of bird lines 64 mya and dispersed throughout the world.
Hoatzins do not fly well, so for those
who argue that opisthocomids evolved in one land and flew over vast oceans to
populate different lands will have a problem. In my opinion, hoatzins dispersed
early on when continents and land bridges were much closer and not later on in
the Cenozoic when huge oceans separated the current continental
distributions.
 |
| Hoatzin chick with forelimb claws |
 |
| A horned screamer’s claws |
 |
| A duck with forelimb claws |
It is true that in the developing embryo
of a bird and human, the pallium (layers of grey and white matter covering the
upper surface of the cerebrum in vertebrates) in birds is derived from the
lower surface of the roof of the fetal forebrain, while in mammals, including
humans, the neocortex is derived from the developing fetal brain’s upper
surface. That is an extrapolation of convergent evolution and not associated
gene evolution. The neocortex along with the archicortex and paleocortex, in
which all three are cortical parts of the cerebral cortex, are involved in
higher functions such as sensory perception, generation of motor commands,
spatial reasoning, conscious thought, and vocally lingual communication.
However, in birds and mammals and again,
this includes humans, the same genes are responsible for the proteins in the
developing embryonic forebrain that produce the proliferation and arrangement
of neurons. It just so happens that this occurs on the lower surface of the
pallium in birds and on the upper surface with mammals.
 |
| Artist: Tony Angell a crow’s brain |
In thinking, requisitions required for
the brain to interpret and interpolate are the chemical messages, the
electrical pathways and the mechanical apertures to produce them. Both the bird
brain and mammalian brain utilize the same genes, neural electrical
transmitters, enzymes and hormones as messenger carriers such as adrenalin,
corticosterone, dopamine, opioids and the FoxP2 gene that is critical to
stimulating nerve development and connectivity in social animals for language
and song learning. Just as in humans, birds can reconsider sensory information
by rewiring neural circuits that loop between the thalamus and pallium.
 |
| Artist: Tony Angell neural looping |
Wild crows play and play a lot, even
adults. It appears to bolster their insight as a motivation to explore, solve
problems, assess situations, sharpen their ability to think of the past to plan
for the future, or simply to frolic.
In Washington State, crows were seen
flying in formation on a cold winter day only to break formation and dive bomb
to the top of a water tower where warm air was rising from it. As the warm air
rose, the crows would spread their wings allowing the rising warm air currents
to glide them upwards until the rising warm air was saturated enough with the
surrounding cold air that it would no longer support any more lift. From there
the crows dove back down repeatedly to the top of the tower to rise but once
again.
A crow at play will pick up a stick and
poke another with it, until the one being poked grabs the stick away from the
poker, switching roles in turning the poked into the poker. On the tips of
branches or wires tied to branches, crows have been seen grabbing the branches
and wires with their beaks or feet then head knob until they begin bouncing up
and down. Ravens have been seen bobsledding down snowy hillsides either on
their backs or belly and once down the hill fly back up to do it again.
 |
| Artist: Tony Angell Ravens wind surfing |
These forms of play activity builds,
re-circuits and refines synapses in the bird’s brain developing required social
skills. This neurobiology of play, as in mammals rewards brain circuits with
new chemistries. Important regions of the social brain affect certain sections
of the forebrain such as the amygdala, which acquires and retains memories sorting
out the pleasant from unpleasant emotional experiences and the septum which
integrates the information from those memories and the hippocampus that affects
social bonding.
As far as emotional sympathy or even
sadness goes, the bird’s brain can also analyze situations calling for these
symptoms. Harpy eagles of Brazil are monogamous and mate for life. When one
loses a mate, they will mourn by incessantly crying out in screeching. While
back to crows, these birds have been seen lining up to view a fallen associate
as the victim of car hit. It’s almost like in a human’s funeral, where
congregants line up to view the deceased at a funeral for the last time. Crows
have even been seen helping an injured crow move by two other crows holding the
injured one up from both sides.
By no means did I go over all the
extinct bird-like and bird fossils. To see a more detailed listing, click on
the link below:
In
Summation:
So, down through the Archosauromorph line from crocodilians
through dinosaurs and finally, to birds we’ve covered this evolution through
the study of fossils.
Fossils reveal that dinosaurs originated
in what is now South America, and geologically soon afterwards first diverged
into sauropodomorphs, then ornithischians and theropods before dispersing
across the Triassic world more than 220 million years ago.
Fossils reveal a lot; much more so than
as simply a species identifier. Youth variations versus adults, what they ate,
whether they chewed or swallowed food whole, how they digested what they ate,
body coverings in skin, scales and feathering, environmental conditions they
existed in, temperature ranges and how they nested; all are revealed in
fossils.
 |
| T. rex jaws infected by trichomonas (arrows) |
 |
| Artist Chris Glen T. rex with trichomonas |
A T.
rex skull fossil shows structurally
different holes bored through the jaw and skull. These are not tooth puncture
wounds and appear to be holes bored by a parasite and in particular the
parasite, Trichomonas that affects
birds today.
 |
| Artist: Jan-Ake Winqvist Maiasaura |
Fossils also give us a window to peer
into dinosaur parental care. ‘Two Medicine Formation’s Egg Mountain in northern
Montana displays parental care in a nesting colony site of the dinosaur, Maiasaura (My-uh-sawr-uh). Numerous nest
sites, eggs and young were found. The eggs were arranged in dug out earthen
nests that were an adult’s body length in distance from each other. The baby
fossils displayed skeletons that still were composed of more cartilaginous
material than bone tissue, too much so that they could not yet walk. This
alludes to the fact of altricial conditions, so parental care would’ve been
necessary for young survival. Also, in the different stages of the baby and
young ages, Maiasaura youth grew
extremely fast as comparable to bird hatchlings and fledglings.
Of course fossils give details of
anatomical structuring. The dinosaur vertebral column consists of the cervical
(neck), dorsal (back), sacral (hips), and caudal (tail) vertebrae. Saurischian fossils
tell us that dinosaur vertebrae sometimes possessed features known as
pleurocoels (hollow depressions on the lateral portions of the vertebrae) that
are perforated creating an entrance into the air chambers within the vertebrae,
serving the dinosaur in decreased bone weight without sacrificing strength. Pleurocoels
were filled with air sacs in furthering weight deduction.
 |
| Pneumatic pleurocels |
As the largest known land vertebrates,
sauropod dinosaurs’ pleurocoels and air sacs may have reduced the animal’s weight
by over a ton, a handy evolutionary adaption in animals that grew to over 30
meters in length. In many hadrosaur and theropod dinosaurs, the caudal
vertebrae were reinforced by ossified tendons.
Going all the way back to the avialans 125
mya, Sapeornis lacked teeth as an
adult in the lower jaw, while Jeholornis
was in the process of reducing teeth; changing the typical dinosaur snout into
an atypical bird beak.
Latter enantiornitheans were trending
towards stronger flight just like the euornithean progenitors of modern birds.
Through convergent evolution, the enantiornithean, Mirarce (Mir-r-say), though totally separate from the euornithean
line of modern birds had evolved highly refined adaptations for advanced
flight.
The fossil of Mirarce shows it still retained more primitive enantiornithean
anatomical features, but also had evolved a more deeply keeled sternum for
stronger flight muscles to attach to and unlike its enantiornithean relatives
in possessing a U-shaped furcula (wishbone), its furcula was more V-shaped as
in euornitheans and modern birds. However, by the time the K-Pg extinction
came, all enantiornitheans had become extinct, even Mirarce who lived 76-74.1 mya.
 |
| Artist: Brian Engh Mirarce atop Utahceratops |
Once you put all that is known of
vertebrate fossils, it becomes a page-by-page book with chapters leading into
the next building up a story through time in how little-by-little, bit-by-bit
one ancestral extinct animal evolved into a more modern one due to evidenced
environmental change and natural selection pressures.
The most costly mobility expense in
energy is flight. We just touched base, but it is truly an exciting event in
seeing how ornithuromorph fossils had already provided the pathway for modern
birds to inherit a specialized digestive system that is lightweight, efficient
and capable of sustaining the energetic demands of powered flight.
The most unique inherited feature of
birds is in having feathers that enabled flight, provides insulation,
camouflage protection and visual communication. Modified feathers aid in
swimming, sound production, water repellence, tactile sensation, hearing, and
body support. Theropdal adaptations for flight include fusion and reinforcement
of lightweight bones and the presence of a keeled sternum in supporting flight
muscles, homeothermy (warm-bloodededness), produce external eggs, and
demonstrate complex parental and reproductive behaviors. Other reptilian and
most likely theropodal shared features include nucleated red blood cells, a
single middle ear bone, and a single occipital condyle on the back of the
skull. Birds uniquely possess vocalization usage to mediate social
interactions, have distinctive bills/feet and are able to detect and react to
magnetism.
 |
| Credit: Ikononet. com Distinctive bird beaks and feet |
In ‘Et Tunc Nulla Erat VII’, from large
toads to pterosaurs to even early day mammals, we mentioned how dinosaurs were
preyed upon by other animals, but dinosaurs surely had other animals, besides
other dinosaurs on their menu lists as well, as is the case in the picture
below.
 |
| Artist: Luis V. Rey Carcharodontosaurs/Ornithocheirus |
I’ll leave you with Spain’s 129.4-126.3 million-year-old
‘La Huérguina Formation’s newly 2018 discovered fossil of a recently hatched
juvenile enantiornithean. Due to the fossil’s exceptional detail, researchers
know it was an enantiornithean, but due to its very recent discovery, have not
as yet assigned it to a genus or species nomenclature.
 |
| The enantiornithean hatchling |
For those that still don’t think birds
came from dinosaurs…have ya ever stared back into the eyes a shoebill stork…
 |
| The Shoebill Stork |
For the next ‘Et Tunc Nulla Erat’
series, we’re gonna go back in time, revisit and pick up the synapsids that
eventually lead to mammals.
Happy Birding
BJA
03/31/2019
No comments:
Post a Comment