Et tunc nulla erat V
(And Then There Was)
The
Road to Archosaurs:
Before we get started with archosaurs,
as we did in the last article, we’ll do a little bit more briefing on
cladograms. First off, remember anytime an animal group name ends in: ‘id’ in
technical reporting, it is referring to the group’s Family identification, which
properly ends in: ‘idae’ in technical nomenclature. Properly named Orders end in: ‘ia’ ‘ria’, ‘es’ or ‘era’ for this writing. In the taxonomic advent of new fields of study,
such as phylogenetics, cladistics, and systematics, the Linnaean structure has
progressed to group rankings of modern biological classification based on the
evolutionary relationships between organisms, both living and extinct. In
cladistic nomenclature, the Order suffix usually ends in formes meaning,
‘having the form of’. The two graphs below help explain descending lineage and where
speciation diversifies from the ancestral lineage.


Monophyletic clades display a taxon or group of organisms consisting of a basal common ancestor and all its descendants. Paraphyletic clades, with ‘para’ meaning ‘almost’ or ‘nearly’ refers to a common ancestor, but excludes one or some of the monophyletic groups. Polyphyletic clades display convergent evolution where common features of a group were not inherited by a common ancestor.


In Et Tunc V under Diapsida, we discussed Archosauromorpha, Archosauriformes and touched on ‘archosaurs’. A breather you deserve. It’s true there is a lot of similar technical jargon to keep pace with here, but please bear with me. Except for mammals and lizards, what most current vertebrate land animals the world over have in common today are in being progeny to ‘archo’- animal groupings.


Monophyletic clades display a taxon or group of organisms consisting of a basal common ancestor and all its descendants. Paraphyletic clades, with ‘para’ meaning ‘almost’ or ‘nearly’ refers to a common ancestor, but excludes one or some of the monophyletic groups. Polyphyletic clades display convergent evolution where common features of a group were not inherited by a common ancestor.


In Et Tunc V under Diapsida, we discussed Archosauromorpha, Archosauriformes and touched on ‘archosaurs’. A breather you deserve. It’s true there is a lot of similar technical jargon to keep pace with here, but please bear with me. Except for mammals and lizards, what most current vertebrate land animals the world over have in common today are in being progeny to ‘archo’- animal groupings.
Archosauromorpha, or
archosauromorph(s) is an infraclass containing all the archosaur clades that were
diapsid reptilians first appearing in the Permian Period 255 mya from Sauria ancestry. Archosauriformes, or archosauriform(s) is a clade that branch evolved from
archosauromorph ancestry, while the true archosaur groups derived from the
archosauriform, Archosauria Order
common ancestor.
So, to make it simple, from common
ancestor to predecessor the line goes: Archosauromporpha
→ Archosauriformes → Archosauria → Archosaurs.
This points out the fact that archosaurs
are archosauriforms and archosauromorphs, but archosauromorphs are not
archosauriforms or true archosaurs and as well, archosauriforms are not true archosaurs.
To add to the admixture, now, archosauromorph
groups have been placed under the new clade, Crocopoda (Phonetics: Croc-o-poda) meaning crocodile foot.

Along with Archosauriformes, also included in the archosauromorph infraclass
and Crocopoda clade are: Rhynchosauria (Phonetics:
Rink-o-sor-ee-ah) and Trilophosauria
(Phonetics: Tril-o-pho-sore-ee-ah). Rynchosaurs have their own clade, where
trilophosaurs now have been put in the newly grouped Allokotosauria (Phonetics: Al-low-kot-o-saw-ria) clade. The clade
name is derived from Greek meaning ‘strange reptiles’ in reference to their peculiar
features as herbivores. Azendohsauridae (Phonetics: Ah-zen-doe-saw-rus)
is also in the Allokotosauria clade
consisting of two herbivorous species. Choristodera
(Phonetics: Core-is-toe-dira), once considered an archosaur, is now determined to
be a sister group to archosauromorphs in both sharing the Sauria clade as their common ancestor.

 |
| The rhynchosaur: Scaphonyx |
 |
| scaphonyx head |
Rhynchosaurs, which looked like a
hammerheaded pig head with a wedged skull that was wider than long, were
archosauromorph herbivores. The trilophosaurs were beaked herbivorous archosauromorphs,
while choristoderes were semi-aquatic resembling modern gavials with a long
narrow snout and were primarily piscivorous, but most likely snatched up any
small aquatic prey with their sharp conical teeth.
Prolacertiformes (Phonetics: Pro-lah-sir-tee-for-mees) meaning ‘before lizard forms’, were long necked diapsids such as Tanystropheus (Phonetics: Tan-ee-stro-fee-us) protosaurids living during the Late Permian into the Triassic. I mention this here,
 for the genus, Prolacerta, meaning ‘before lizard’ is represented by only one species, Prolacerta broomi. But Prolacertiformes was named after Prolacerta that is now considered a
sister taxon to Archosauriformes due
to the most recent phylogenetic analyses. This means Prolacertiformes is also archosauromorph derived, although their
phenotypes appear to have converged or reverted through convergent evolution to
be the same. These like appearances may not have been inherited from a common
ancestor. Therefore, Prolacerta is
now the single species in Prolacertiformes
while the other animal groups formerly listed there have now been moved to the
order, Protosauria, but all are still
considered to be archosauromorphs. Boreopricea
funerea is a single species so far found in the genus and at only
10cm/3.94in in body length, is considered a small protorothyrid related to the
larger protorosaurs, such as Pamelaria
and Jaxtasuchus.
for the genus, Prolacerta, meaning ‘before lizard’ is represented by only one species, Prolacerta broomi. But Prolacertiformes was named after Prolacerta that is now considered a
sister taxon to Archosauriformes due
to the most recent phylogenetic analyses. This means Prolacertiformes is also archosauromorph derived, although their
phenotypes appear to have converged or reverted through convergent evolution to
be the same. These like appearances may not have been inherited from a common
ancestor. Therefore, Prolacerta is
now the single species in Prolacertiformes
while the other animal groups formerly listed there have now been moved to the
order, Protosauria, but all are still
considered to be archosauromorphs. Boreopricea
funerea is a single species so far found in the genus and at only
10cm/3.94in in body length, is considered a small protorothyrid related to the
larger protorosaurs, such as Pamelaria
and Jaxtasuchus.
 |
| The choristodere: Champosaurus |
Prolacertiformes (Phonetics: Pro-lah-sir-tee-for-mees) meaning ‘before lizard forms’, were long necked diapsids such as Tanystropheus (Phonetics: Tan-ee-stro-fee-us) protosaurids living during the Late Permian into the Triassic. I mention this here,

 |
| Protorosaurs |
We will not elaborate any further on the archosauromorph groups except for those that led to the Archosauriformes clades as the others all became extinct leading to a dead end along the evolutionary line. Except for choristoderes, the rhynchosaur and trilophosaur species all died out by the end of the Triassic extinction; choristoderes lived a long span from the Middle Jurassic to the Miocene, a span from 165 to 20 mya.
 |
| Trilophosaurus buettneri fossil Note:beak, flattened cheek teeth |
 |
| Protorosaurus fossil |
From the mid-Devonian onwards, land
witnessed a widespread diversification of trees and shrubs and herbivore
trending archosauromorph protorothyrids took advantage of it through natural
selection in evolving shifts. To access higher edible plant parts, these
animals also were rather large with longer hind limbs in so far as being the
first quadruped to show biped abilities in rearing on hind legs to reach higher
up in treetops.
Archosauromorphs/Archosauriforms:

On the path to archosauromorphs, the clade, Sauria (Phonetics: Saw-ree-ah) is the first bump in the road encountered. Atopodentatus (Phonetics: Ah-top-oden-ta-tus) was an oddball saurian and thus far, is the earliest marine herbivore found. Being semiaquatic while swimming the Middle Triassic sea shorelines 240 mya, Atopodentatus with its hammerhead rimmed with small cutting teeth would chisel out the seafloor foraging for algae.
The first fossil finds misinterpreted
that the upper jaw had a centered vertical split where the teeth fitted
together in a zipper-like fashion for filtering. Later fossil finds disproved
this bearing out the hammer styled head. Although in either case the head was
indeed odd, the body was already showing trending archosauromorph features with
robust appendages, longer hind limbs, hip arrangement and in the morphology of
the cervical vertebrae.
These two archosauromorphs survived the ‘Great Dying’ at the Permian/Triassic border. Due to the ‘Lilliputian Effect’ where post extinction smaller specimens are favored, the long necked Czatkowiella (Phonetics: Zat-ko-wee-el-ah) was small at 1.5m/4.9ft. Protorosuarus (Phonetics: Pro-toe-ra-sore-us) was almost double that reaching a length of 2.64m/8.67ft, but was considered small for what was to evolve as predecessors. At times these two basal archosauromorphs are described as semi-aquatic, most likely though, they were fully terrestrial.

On the path to archosauromorphs, the clade, Sauria (Phonetics: Saw-ree-ah) is the first bump in the road encountered. Atopodentatus (Phonetics: Ah-top-oden-ta-tus) was an oddball saurian and thus far, is the earliest marine herbivore found. Being semiaquatic while swimming the Middle Triassic sea shorelines 240 mya, Atopodentatus with its hammerhead rimmed with small cutting teeth would chisel out the seafloor foraging for algae.
 |
| Atopodentatus |
These two archosauromorphs survived the ‘Great Dying’ at the Permian/Triassic border. Due to the ‘Lilliputian Effect’ where post extinction smaller specimens are favored, the long necked Czatkowiella (Phonetics: Zat-ko-wee-el-ah) was small at 1.5m/4.9ft. Protorosuarus (Phonetics: Pro-toe-ra-sore-us) was almost double that reaching a length of 2.64m/8.67ft, but was considered small for what was to evolve as predecessors. At times these two basal archosauromorphs are described as semi-aquatic, most likely though, they were fully terrestrial.
 |
| Depranosuarus (Note: clawed finger and prehensile tail tip) |
 |
| Depranosaurus |
 |
| Tanystropheus |
 |
| Tanystropheus, semiaquatic |
Eventually around 260 mya ago, Archosauriformes refined the archosauromorph’s line of evolvement. Archosauriformes still retained the Archosauromorpha thecodont socketed teeth, but were more derived diapsid reptiles. In skull morphology, Archosauriformes species had strongly downturned premaxillae, had a pair of plate-like bones known as septomaillae and the nasals were restricted to the dorsolateral surface of the snout possessing an extensive suture with the maxilla extending from anteroventral (in relation to underside front of the head) to posterodorsal (situated posteriorly on the underside of the head). Also, the hind limbs had extended even more physiologically on average than other archosauromorph groups. There are other varying characteristic traits, but to laundry list it here would become too boring for this treatise.
During the Triassic 250 mya ago, a branch
of archosauriforms evolved into the clade members of, Crurotarsi (Phonetics: Crew-row-tar-see). With the Latin word, crus combined with the Greek word, tarsos, the clade name refers to
specialized articulation between the fibula of the limb and calcaneus of the
heel. This is witnessed in all archosaurs under Archosauria, as all extinct crocodilians, extant crocodilians, all
birds (which are avian dinosaurs), non-avian dinosaurs, pterosaurs and the
extinct phytosaurs possess the arrangement or a version of it.

The archosaurian phlyogeny split into two group clades. Originally named the Pseudosuchia and Ornithosuchia, they have currently been replaced since 1990 with Crurotarsi for the crocodilimorphs and Avemetatarsalia, or sometimes Ornithodira with both referring to all crown archosaur groups related closer to dinosaurs rather than crocodilians. Just remember though, both sets such as Pseudosuchia opposing Crurotarsi are still both used singly and interchangeably as I have done as well in this treatise.
Archosaurs are (I said ‘are’ instead of ‘were’ for archosaurs are still with us in the form of extant crocodilians and birds) diapsid tetrapods, but in addition of distinguishing them from all other diapsids, are an extra pair of openings in front of the eye sockets known as antorbital fenestrae. Where crocodilians have lost this anatomical process, birds still retain it, while in other later evolved archosaurs the fenestrae digressed into a small depression known as the antorbital fossa. The antorbital fossa is a basal depression including the antorbital fenestra, which is a skull opening and as well includes the surrounding fossa. Later archosaur trends will be to close the fenestra opening, yet retain pneumaticity in the antorbital region.
In general, archosaurs anatomically had
larger hind limbs than the forelimbs, a more narrow longer snout as compared to
the ancestral reptiliomorphs and the modified ankle that gave greater ease in
pivoting.
By the time the true archosaurs came
around 245 mya, there was a great diversification in speciation with increasing
sizes. There were many herbivores and carnivores occupying all ecological
niches from being fully terrestrial, semi-aquatic and to being fully aquatic.
This was due to the archosaur line and in so doing the Triassic became the ‘Age
of Reptiles’.
As witnessed through taxonomic phylogeny and putative detection of several proteins, hemoglobin, etcetera in extant genera DNA gene sequencing, the clade chart below relays this burst of diversification.
As witnessed through taxonomic phylogeny and putative detection of several proteins, hemoglobin, etcetera in extant genera DNA gene sequencing, the clade chart below relays this burst of diversification.


From archosauromorphs evolved the true archosaurs. Archosaurs came from the Archosauriformes clade, Crurotarsi (phonetics: Crew-row-tar-sigh). Crurotarsans are so named due to the specialized articulation between the crus (Latin: bone between fibula/tibia and tarsus) and tarsus (Greek: tarsos) bones of the foot. These are located between the fibula’s ankle end and the calcaneum (heel bone). The term crurotarsan literally means ‘cross ankles’. Extant crocodilians retain vestige features of this unique foot arrangement between the fibula and heel called crurotarsal, while dinosaurs held and hold a revised ankle joint arrangement known as mesotarsal.
Below is a cladogram of crurotarsans and
we will only elaborate a bit through the crurotarsans (archosaurs) to phytosaurs,
euparkians, aetosaurs and rauisuchians.

Phytosaurs (extinct), aetosaurs
(extinct), crocodilians, ornithosuchians (extinct), rauisuchians (extinct), non-avian
dinosaurs (extinct) and birds are all descendants of the archosaurimorph node
clade, Crurotarsi enlisting all the archosaurian
forms.
Showing up a little before 251 mya in
the Triassic, crurotarsans are the most primitive of archosaur phytosaurs and
suchians (the crocodile line).
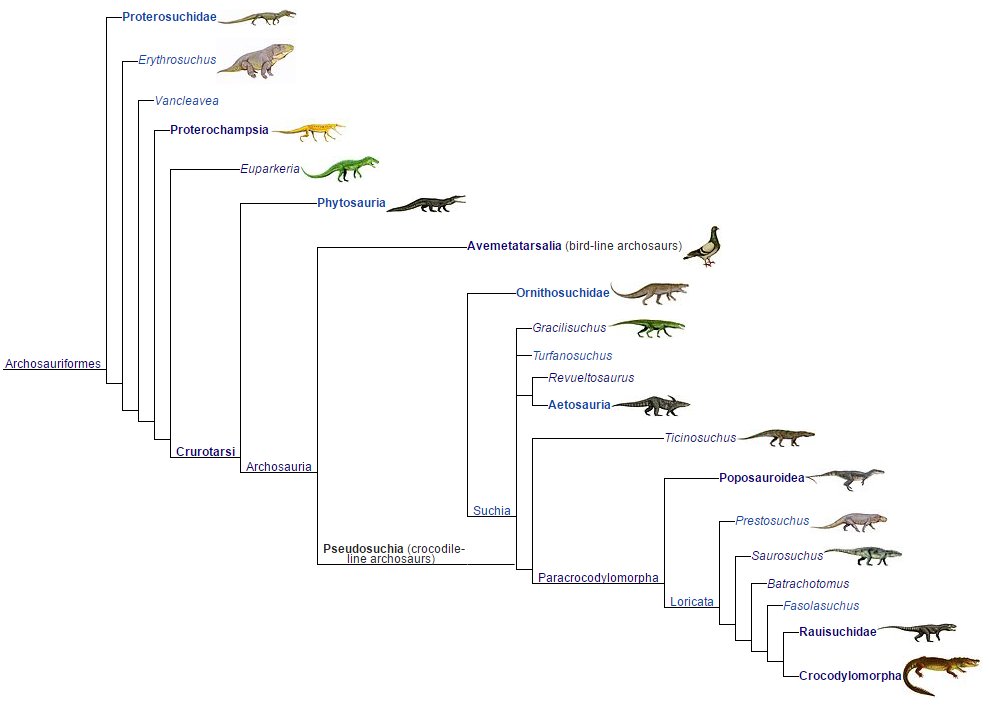
The archosauriforms listed under the clade, Poposauroidea (Phonetics: Pop-o-sore-oid-e-ah) are basal to the Ctenosauriscidae (Phonetics: Ten-o-sore-is-see-die) clade. These clade names front crurotarsan archosaur species such as, Poposaurus and the sail-backed, Ctenosauriscus, but not rauisuchians or aetosaurs, so therefore are used to model the earliest crocodilian lineages.

One of the earliest of archosauriforms was Xilousuchus (Phonetics: Zee-lew-sue-kuss) arising in the Early Triassic of the Olenekian Stage 250-247 mya. Originally classed as a protosuchid, but after careful phylogenic analysis in 2011 by the famed paleontologist, Sterling Nesbitt, the animal was relocated back further in time as one of the first of the primitive crurotarsan archosaurs.
Not looking much like anything that
would descend from this ancient archosaur, (as all sphenodonts, crocodilians,
dinosaurs, extant reptiles and birds eventually did), Xilousuchus had elongated vertebra neural spines that supported a
sail. This primitive archosaur’s sail is convergent evolution as the synapsid
family of dimetrodons supported sails, as well as the theropod dinosaur, Spinosaurus (Phonetics:
Spine-o-sore-us). All were from varying time periods and at best were remotely
related.
 |
| Xilosuchus |
Xilousuchus was a
quadruped, but it possessed longer hind limbs as compared to the forelimbs. Its
sail would have projected the animal’s center of gravity too far forward to
have actually made it bipedal, but it would have allowed the animal to stand on
its hind limbs.
 |
| Arizonasaurus |
 |
| Ctenosauriscus |
 |
| Euparkeria skeletal strucure |
 |
| Euparkeria |
 |
| Sclerotic ring |
Euparkeria was truly one of the first
tetrapods to being cursorial in that they were adapted for speed. The Euparkeria line is also now viewed to
have been basal to all archosaurians.
 |
| Longisquama fossil |
 |
| Longisquama with flesh |
Phytosaurs:

As a sister taxon to crurotarsans, the archosauriform phytosaurs were members of the order, Phytosauria (Phonetics: Fi-toe-sore-e-ah). Phytosaurs first evolved in the latter part of the Middle Triassic 228 mya but all were extinct by the end of the Triassic 199.6 mya. Evolutionary relationships dictate that phytosaurs evolved before the split between suchian crocodilian and bird-line archosaurs.
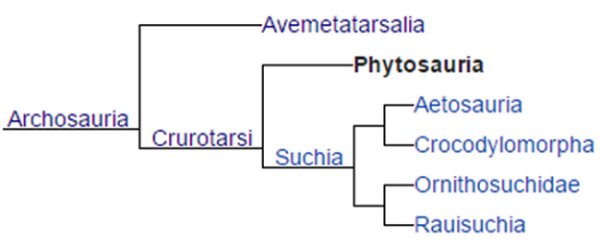
As a sister taxon to crurotarsans, the archosauriform phytosaurs were members of the order, Phytosauria (Phonetics: Fi-toe-sore-e-ah). Phytosaurs first evolved in the latter part of the Middle Triassic 228 mya but all were extinct by the end of the Triassic 199.6 mya. Evolutionary relationships dictate that phytosaurs evolved before the split between suchian crocodilian and bird-line archosaurs.
Phytosaurs were semi-aquatic inhabiting
the inland freshwater shorelines during the Late Triassic. The name phytosaur
is a misnomer for it means, ‘plant eater’ in which the paleontologists in
discovering the first fossils thought they were. Later fossil finds exhibiting
sharp conical serrated teeth bore out that they were indeed predators.
 |
| The phytosaur: Rutiodon |
Anatomically resembling crocodiles and gavials/gharials, the differences are in having a much more primitive ankle structure than crocodilians, were more heavily armored with heavy bony body scutes, nostrils were placed just below the eyes where crocodilian nostrils are located near the end of the snout and the belly was reinforced with a dense arrangement of abdominal ribs known as gastralia. With various snout crests, dolichorostral ‘long snouted’ types had a long, slender snout as gavials, where brachyrostral ‘short snouted’ forms are the opposite, having a massive, broad snout more like alligators. Phytosaurs were a successful group of animals and had a global distribution.
 |
| The phytosaur: Redondasaurus bermani |
The crocodile body plan and type livelihood have been around a lot longer than crocodiles have. Phytosaurs took over the lie in wait ambush aquatic bank strategy with a crocodilian morphology from the stereospondyl trematosauroids. Crocodilians did the same in filling this ecological niche once phytosaurs exited life.
The difference was that phytosaurs as a
sister taxon are more closely related to crocodilians where both diapsid groups
filled the amphibian trematosauroid cranny through convergent evolution. Neither
group had a direct ancestral lineage with trematosauroids.
When phytosaurs died out, the
crocodylomorphs rapidly evolved almost into the same phytosaurian form once
filling the niche phytosaurs had abandoned.
 |
| Prionosuchus restored skeletal fossil |
 |
| The Temnospondyl, Prionosuchus |
To add a note here, in the photos above,
most at first glance would surmise that it represents some crocodilian like a gavial,
but it is not. The restoration photos depict a temnospondyl salamander. Its
name is Prionosuchus (Phonetics:
Pree-on-o-sook-us), a salamander that lived in the Permian 270 mya, long before
the first archosauromorph came into being. It just goes to show that convergent
evolution can elude discerning polyphyletic genetics.
Aetosaurs:

The order Aetosauria (Phonetics: e-uh-toe-sor-e-uh) were archosaurs that, although fully terrestrial, were close early relatives to semi-aquatic crocodiles where both groups derived from early pseudosuchian archosaurs. Dentition shows that aetosaurs were strict herbivores, but in the aetosaur genus, Saurosuchus (Phonetics: Sar-o-sook-us), with backward pointing sharp teeth, it may have been omnivorous. Aetosaurs had a pillar erect posture, swaying to evidence they were agile and quick in locomotion.

The order Aetosauria (Phonetics: e-uh-toe-sor-e-uh) were archosaurs that, although fully terrestrial, were close early relatives to semi-aquatic crocodiles where both groups derived from early pseudosuchian archosaurs. Dentition shows that aetosaurs were strict herbivores, but in the aetosaur genus, Saurosuchus (Phonetics: Sar-o-sook-us), with backward pointing sharp teeth, it may have been omnivorous. Aetosaurs had a pillar erect posture, swaying to evidence they were agile and quick in locomotion.
 |
| Revueltosaurus |
The pseudosuchian suchian, Revueltosaurus (Phonetics:
Ray-veil-toe-sar-us) with primitive morphological and anatomical features akin
to aetosaurs may have been a direct ancestor.
Occurring in and going extinct by the
end of the Late Triassic (235-208.5 mya), aetosaurs with the name meaning,
‘eagle lizard’ kind of reminds one more of an anklyosaurus dinosaur
anatomically, but, as an archosaur, were one of the closest relatives to
crocodilians. All aetosaur species have only been found in Late Triassic
sediment layers when all continents were connected during Pangaea.

On average, aetosaurs were 3m/9.9ft long
with species ranging from 1m/3.3ft to 5m/16.5ft. Along with an upturned snout,
anatomical shorter forelimbs and stout clawed feet features show they were adapted to
digging. As modern day crocodilians, aetosaurs built complex and elaborate
nests by honing out rock into horseshoe or circular depressions lipped with
high ridges. Many Desmatosuchus
(Phonetics: Dez-mat-o-such-us), a horned aetosaur footprints were found in
northeastern Italy all around and near the nest sites.
 |
| Newly discovered aetosaur: Gorgetosaurus |
Please note: except for Xilosuchus, in which the ‘suchus’
portion in the name is ‘sue-kuss’, all other ‘suchus’ nomenclature are pronounced
‘such-us’ or ‘sook-us’. I chose ‘such-us’, but most certainly ‘sook-us’ can be used interchangeably.
Rauisuchians:


Rauisuchia (Phonetics: Ral-ee-sue-chee-ah)
occurred during the Triassic and ranged in size from 3.9m/13ft to 6.1m/20ft.
Along with aetosaurs, rauisuchians in having a hip socket face downward in
forming a shelf bone for the femur to connect underneath, had a pillar erect
gait. They also had a well-defined rotary joint between the astragalus (talus)
and calcaneum that was very similar to crocodilian ankles. All of the species
possessed a thickening, or a ridge or rugosity on the anterolateral surface of
the ilium positioned immediately above the acetabulum.

Rauisuchian hind limbs were much longer
than the forelimbs, but except for Postosuchus,
who was bipedal with much reduced forelimbs and foot digits, the rest were
quadrupeds. Rauisuchians had a pillar erect stance much like aetosaurs.
 |
| The rauisuchian, Fasolasuchus |
Most Rauisuchian skulls were heavily built that bore recurved dagger-like teeth. Species ranging from 4m/12ft to 6m/20ft, Rauisuchians were carnivorous and must have been an apex predator, for only after they had died out did dinosaurs begin to make their rise.
 |
| Postosuchus fossil |
Postosuchus (Phonetics: Pos-toe-such-us), from the Late Triassic 221-203 mya, it was a typical rauisuchian, with a massively built skull that was narrowly constructed anteriorly while extending deep and wide posteriorly. The skull was riddled with fenestrae to alleviate weight and act as muscle anchoring. It also possessed osteoderms (thick plated scales) than ran the course of the dorsal side from neck to tail.
 |
| Postosuchus in the flesh |
Standing at 1.2m/3.9ft in height, Postosuchus measured 4m/13ft long and weighed ~ 275kg/255lbs. With the forelimbs around 64% smaller than the hind limbs, it surely most likely preferred bipedal locomotion for accuracy and speed, although it could walk in tetrapodal fashion as well. All four limbs were situated directly under the body, but over osteological aspects, were structured for a leaning gait rather than a straight up posture.
With dagger-like teeth, this apex
predator was surely the scourge for herbivores sharing its warm tropical
ecosystem of what was to become the United States where Postosuchus fossil remains have only been found.
Crocodylomorphs/Crocodyliforms:
Just as we did for archosaurs, we’re
going to look at the crocodylomorphs that lead to crocodyliforms that gave rise
to the true crocodiles in crocodilians, therefore: Crocodylomorpha → Crocodyliformes → Crocodilians.
From the main primitive common ancestor to crocodylomorphs was Suchia (Such-ee-ah) and through its many branchings led paraphyletically to paracrocodylomorphs that became a paraphyletic common ancestor to crocodylomorphs.

The Crocodylomorpha
members are the only crurotarsan/pseudosuchian archosaur animal groups that
survived the Triassic/Jurassic extinction, whereas, Crocodyliformes are the only crocodylomorphs that survived beyond
the Early Jurassic. Below is the phylogenetic taxon relationship in dendrogram
form.

An early archosaur occurring during the
Middle Triassic (242-235 mya) belonged to the Paracrocodylomorpha (Ponetics: Pair-ah-crah-co-dill-o-mor-pha)
clade that, along with other archosaur branches, eventually gave rise to
crocodylomorphs. One paracrocodylomorph archosaur belonged to the genus, Batrachotomus (Phonetics:
Ba-trac-o-toe-mus) and was a formidable predator in the Triassic swamps it
roamed reaching 6m/20ft in size. The name comes from Greek with ‘batrach’
referring to ‘frog’ and ‘tome’ to ‘slicing’. This alludes to its cutting edged
teeth and to the amphibians it preyed on in the swampy environs, in particular
the 1.25m/4ft salamander, Mastodonsaurus.
 |
| Batrachotomus |
With Batrachotomus’ upright stance due to hip and joint socket arrangements, later crocodylomorphs would inherit its erect stance with smaller forelimbs than hind limbs making for agile locomotion.
 |
| Gracilisuchus |
Crocodylomorphs also filled about every
niche available evolving from species less than 30.5cm/1ft in the
earliest of crocodylomorphs within the sphenosuchian genus, Gracilisuchus (Phonetics:
Gra-sah-le-such-us), to later ones reaching over 3m/10ft, such as the 231 million
year old bipedal Carnufex (Phonetics:
Car-nu-fex). Carnufex is Latin for ‘butcher’.
 |
| Terrestrisuchus |
Gracilisuchus and the larger,
more derived sphenosuchian crocodylomorph, Terrestrisuchus
(Phonetics: Tuh-res-tri-such-us) at 80cm/2.6ft, were strictly terrestrial
animals. Bearing no resemblance to modern day crocodiles, their thimble legs
were designed for galloping and sprinting. Inherited from these much smaller
earlier crocodylomorphs, modern day crocodiles today exhibit short bursts of
terrain galloping.
Carnufex, a suchian
crocodylomorph was a holy terror of the Late Triassic 231 mya. As a fairly
speedy bipedal predator, it filled a terrain niche and ecological gap in
tropical Pangaea that later was to be filled by theropod dinosaurs. It most
likely preyed on anything it ran down, which included some of the earliest mammals.
 |
| Carnufex carolinensis |
 |
| The herbivore crocodyliform: Chimeaerasuchus |
Chimaerasuchus (Phonetics: Ky-mare-ah-sook-us) was a mesoeucrocodylian
crocodyliform showing up in the Early Cretaceous 115-110 mya. Showing various
teeth structure, which are ziphodont (blade-like), tribodont (bluntly rounded) and
multi-cusped crown teeth, this notosuchian was a strict herbivore. Phylogenetic
analysis has determined that Chimaerasuchus
was a sister group to notosuchian sebecosuchian crocodyliforms.

The Notosuchia (Phonetics: No-toe-such-e-ah) and Neosuchia (Phonetics: Nee-o-such-e-ah) are crocodyliform clades that arose from mesoeucrocodylians splitting the Mesoeucrocodylia common ancestry. Mesoeucrocodylia contains many other crocodyliform groups. It is the paraphyletic clade that neosuchians arose from, eventually leading to eusuchian crocodilians, which is the line we will primarily follow. Below is a much simpler cladogram.

 |
| Simosuchus fossil |
 |
| Simosuchus restoration |
 |
| Aplestosuchus fossil with last meal remains |
Just one more notosuchian we’ll note. An Aplestosuchus (Phonetics: A-pless-toe-sook-us) fossil find was a significant discovery for in its abdominal cavity, it appears its last meal was an unidentified sphagesaurid crocodyliform. We know the sphagesaurid was eaten due to its deteriorated state as compared to the Aplestosuchus remains. This is significant in knowing that one crocodyliform would prey upon another. Due to this the name, Aplestosuchus is Greek derived meaning, ‘filthy gluttonous crocodile’.
 |
| Aplestosuchus with prey |
Where most of crocodyliform ancestry was terrestrial, as just stated, crocodyliforms had also migrated to watery environs. However, not all crocodyliforms were relegated to a semiaquatic lifestyle along riverbanks and shorelines acting as ambush predators waiting for prey to come along.
The neosuchian, Stomatosuchus (Phonetics: Sto-mat-o-sook-us), meaning ‘mouth crocodile’ was perhaps the largest seafaring crocodile of all time with an estimated length of 10m/32.8ft. However, with gracile small conical teeth in the upper jaws, the lower jaws (the mandible) were toothless, but supported a large gular pouch much like a pelican’s. It was not a predator, but a filter feeder known as a planktivore filling the gular pouch with plankton, krill and small fish while patrolling the shallow mangrove shorelines, then squeezing the water out through the teeth trapping its prey and finally swallowing the contents. Stomatosuchus lived during the Late Cretaceous 100-94 mya.
 |
| Stomatosuchus |
Sarchosuchus (Phonetics: Sar-ko-sook-us), was a pholidosaurid crocodyliform living in the Early Cretaceous 112 mya. Known as supercroc, it is the largest member of the crocodylomorph line thus far discovered. It was at least 11m/36.1ft with upper size estimates reaching upwards to 13m/42.7ft. We know this for the osteoderm rings found in one fossil find shows that it was forty-years-old and still growing.
 |
| Sarchosuchus (note bulla snout) |
With 35 teeth on each side of the upper
jaw and 31 on each side of the lower, the jaws were made for seizing and holding
onto prey. For its size and bulk, Sarcosuchus
required larger prey and what was abundant in its day were dinosaurs that it
would ambush and drag into the water to drown then tear and break apart for
swallowing, much like crocodiles and alligators do today.
 |
| Sarchosuchus skull vs. Human & extant croc |
Instead of being semiaquatic, some
crocodyliforms were both semiaquatic and marine venturing into deeper waters,
but still having feet to swim with. Machimosaurus
hugii, a thalattosuchian crocodyliform was just such one of these.
 |
| M. hugii |
 |
| M. rex |
 |
| Dakosaurus |
 Metriorhynchidae (Phonetics:
Met-ree-or-rink-ah-day) were fully aquatic adapted for a total marine life and
although there is no proven fossil finds yet, metriorhynchids might even have
given birth in the waters. Metriorhynchus,
found in Mid-Late Jurassic deposits with a temporal range of 167-155 mya, while
losing scalation for smoother skin, evolved fully whole paddles for limbs while
utilizing a vertically ruddered tail used for sinuous propulsion. It swam the
shallow seas while at times venturing into deeper waters to prey on the gentle 16m/52.5ft
fish, Leedsichthys by taking chunks
of flesh off of the victim. But in deeper waters, Metriorhynchus must’ve been prey to the short-necked pleurodon, Liopleurodon that ate just about
anything that swam its way.
Metriorhynchidae (Phonetics:
Met-ree-or-rink-ah-day) were fully aquatic adapted for a total marine life and
although there is no proven fossil finds yet, metriorhynchids might even have
given birth in the waters. Metriorhynchus,
found in Mid-Late Jurassic deposits with a temporal range of 167-155 mya, while
losing scalation for smoother skin, evolved fully whole paddles for limbs while
utilizing a vertically ruddered tail used for sinuous propulsion. It swam the
shallow seas while at times venturing into deeper waters to prey on the gentle 16m/52.5ft
fish, Leedsichthys by taking chunks
of flesh off of the victim. But in deeper waters, Metriorhynchus must’ve been prey to the short-necked pleurodon, Liopleurodon that ate just about
anything that swam its way.  |
| The gentle giant fish: Leedsichthys |
There were some odd crocodyliforms
either in morphology or habits that one wouldn’t really think of as being in
the crocodilian line. Below are a few.
The Armadillocroc, Armadillosuchus: (Phonetics: Ar-mah-dil-lo-sook-us); Late
Cretaceous 93-84 mya; 2m/6.6ft.
 |
| Armadilliosuchus |
 |
| Armadillosuchus fossil |
 |
| Anatosuchus |
With a duck-like snout, Anatosuchus had a pointed protuberance
at the tip of the snout which has been determined had extra sensory receptors
to root out small fish, frogs and grubs. Once prey was located it snatched them
out with hooked-shaped teeth.

This terrestrial and semiaquatic
notosuchian crocodyliform had limbs for galloping on land and a tail for
swimming in water. On land, it was adapted for speed having an upright
quadrupedal stance with elongate carpals (wrist bones) in relation to the
metacarpals (hand bones). It’s unique dental morphology leads to an omivorous
diet. There were six Araripesuchus
species that most likely evolved from the breakup of Gondwana isolating the
genus in geographical fragmentation. This is known as ecological vicariance.
The Ratcroc, Araripesuchus rattoides: (Phonetics: Uh-rare-ip-uh-sook-us Rat-toi-dees),
Late Cretaceous 100-94 mya; ~ 1m/3.3ft.
 |
| A. rattoides |
With a pair of buck teeth in the lower
jaw this little land crocodyliform used its dentition to root out insects, grubs
and small vertebrates and to dig up plants tubers and roots. It was agile and
quick on land.
The Boarcroc, Kaprosuchus: (Phonetics: Kap-row-sook-us) Late Cretaceous 100-94
mya; 3.3m/10.8ft.
 |
| Kaprosuchus displaying lethal teeth |
This neosuchian, shared several phylogenetic
characteristics with Mahajangasuchus (Phonetics:
Ma-ha-jawn-ga-sook-us). Kaprosuchus
swam swamps, or other slow moving bodies of water, almost fully submerged
stalking water and land animals. On land, it was agile and could display short
bursts of speed. When a land animal was taking a drink on the shore, Kaprosuchus would swim away come on land
and charge its intended victim bowling it over by ramming it with its
reinforced armored snout. Once the victim was down it would use its dagger like
teeth to take hold of its prey to tear or rip it apart.
The Pancakecroc, Laganosuchus: (Phonetics: Lah-gah-no-sook-us), Late Cretaceous 95 mya;
6m/19.7ft.
 |
| Laganosuchus |
Laganosuchus was primarily
piscivorous and with mouth agape, patiently lied in wait for a fish to swim across,
whereupon the jaws would be instantly shut entrapping the victim, then
swallowed. This crocodyliform was dorsally flat from snout to tail, although
the tail most likely did support raised scales on each edge. This stomatosuchid
crocodyliform was very patient lying motionless until the next meal came along
triggering the snapping of the jaws shut.
Below is an excellent ‘National
Geographic’ video explaining the fossil findings and lives of the last five
above mentioned crocodyliforms.
After radiating out in diversity, crocodyliforms
suffered a major biodiversity decline during the boundary between the Jurassic/Cretaceous
in both the marine and terrestrial groups. This is accompanied by elevated
extinction rates in the Late Jurassic, at a near level mass extinction status, severely depressing origination rates in the Early Cretaceous. Sea-level
changes were primarily responsible for this biodiversity decline, both in
marine realm and land species, due to greatly reduced amounts of habitable
shallow marine regions. Also, secondary factors driving biodiversity changes
included perturbations to the carbon and sulfur cycles that, together with
sea-level fluctuations, indicate a prominent role for large-scale tectonic
processes in shaping crocodyliform biodiversity in the Late Jurassic to Early
Cretaceous. There is also evidence for a mediating paleotemperature decreasing effect
on Northern Hemisphere crocodyliform declines during this interval.
Overall, this suggests the fate of
Mesozoic crocodyliforms was coupled more broadly to a combination of
environmental factors with a wider impact on pelagic and shallow marine
ecosystems. Sea-level rise is the principal driving factor in shaping the
evolution of continental shelf biotas. This is not to say that minor additional
ecological factors were also at play across the Jurassic/Cretaceous boundary;
it’s just so far, not evident.
However, several non-marine groups with a
high biodiversity before the boundary extinction, most notably non-sebecid
notosuchians, the marine dyrosaurids and terrestrial sebecids survived
alongside crocodylians.
Nevertheless, the extinctions of these
groups, and other non-marine crocodylomorph taxa were balanced by rapid
radiations of the three surviving clades in the early Paleocene, including
substantial range expansions of marine dyrosaurids and terrestrial alligatoroids
into what is now South America.
We mentioned the methods of how
crocodyliform species could be at home on land just as much in water, for this
high command of environmental domination must be a key factor in it escaping
predation as well as surviving mass extinction periods.
Below is a much generalized cladogram of the archosaur path leading crocodilians.

The next clades to appear were the crocodyliforms that split with one of the two clades, mesoeucrocodylians forming neosuchians.

From neosuchian common ancestry arose the eusuchian clade that gave rise to the order, Crocodilia, the true crocodiles.

Crocodylians:
Below is a much generalized cladogram of the archosaur path leading crocodilians.
To recap in simple cladograms, first
arose crocodylomorphs.

The next clades to appear were the crocodyliforms that split with one of the two clades, mesoeucrocodylians forming neosuchians.

From neosuchian common ancestry arose the eusuchian clade that gave rise to the order, Crocodilia, the true crocodiles.

Crocodylians:
You may bump into your own research with
Crocodilia spelt as Crocodylia. Either way is accepted in
nomenclature.
From the main primitive common ancestor to crocodylomorphs was Suchia (Such-ee-ah) and through its many branchings led paraphyletically to paracrocodylomorphs that became a paraphyletic common ancestor to crocodylomorphs.


Order members of Crocodilia have been around since 83.5 million years. In that time frame, archosaur crocodile morphology hasn’t changed much as exhibited in extant crocodilians of today in comparison to extinct crocodilian species. With that, we call modern day crocodiles/alligators/caiman/gharials as primitive. But, we shouldn’t interpret them as primitive simply because they’ve evolved little change.
From the main primitive common ancestor to crocodylomorphs was Suchia (Such-ee-ah) and through its many branchings led paraphyletically to paracrocodylomorphs that became a paraphyletic common ancestor to crocodylomorphs.
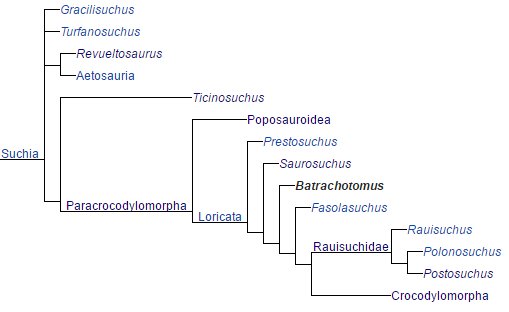
Under Eusuchia (Phonetics: U-such-ee-ah), Crocodilia (Phonetics: Croc-o-dill-ee-ah) arose evolving into the
extant crocodilians we have today. This includes, alligators, caiman, gharials and crocodiles. Below is a clad of all extant crocodilians.

Order members of Crocodilia have been around since 83.5 million years. In that time frame, archosaur crocodile morphology hasn’t changed much as exhibited in extant crocodilians of today in comparison to extinct crocodilian species. With that, we call modern day crocodiles/alligators/caiman/gharials as primitive. But, we shouldn’t interpret them as primitive simply because they’ve evolved little change.
Sharks have been around in their current
general form a lot longer than crocodiles have in their primitive forms. As
geologically spoken in timelines, the Crocodilia
line has not been that long, originating 45 million years ago; the Silurian jaw
explosion on the other hand made its appearance 442 mya, in which ‘Jaws the
Shark’ came soon after.
This should not be an area of question
marks, for evolution works through natural selection of a particular
environment. If that habitat changes, those mutations that were once
detrimental suddenly become an advantage stimulating genomic change and
speciation through natural selection. If environmental stresses do not have
enough input to affect required changes, then the species will survive as it
always has in physiological and morphological survival strategies.
It’s useful in knowing that in the crocodiles’
embryonic stage, unlike other reptiles, develops a four-chambered heart until a
bypass further develops in latter fetal development making it functionally
three chambered; thus evolving from homeotherms into ectotherms. Birds also bypass the heart chamber during embryonic development, but remain warm-blooded.
Too many people consider being ‘cold
blooded’ as an inferior archaic state, for after all, it was the primitive
condition. But in crocodiles it is derived as a reversion to the primitive
condition, because being ‘cold blooded’ is superior in some cases. Crocodiles
actually came from warm blooded terrestrial ancestors.

Susisuchus (Phonetics: Su-see-sook-us) is a neosuchian crocodyliform that lies just outside the clade Eusuchia, but is associated with the transition between basal and advanced neosuchians and the rise of early eusuchians.
As recently as the Eocene in the middle of the Paleogene Period (formerly the Tertiary Period) 56-33.9 mya, there was a crocodilian that lived fully on land and ran. Pristichampsus (Phonetics: Pris-stee-champ-sus) had long cursorial legs adapted for running. This was an advantage to being fully terrestrial and on its topside, the jaws were equipped with ziphodont teeth that were characteristic of land predators to grab and tear prey rather than to simply hold onto prey and drown them as aquatic crocodilians do.
This 3m/9.9ft crocodilian had longer legs than its contemporary semiaquatic crocodilian cousins and in addition, Pristichampsus’s toe ends were hoofed for better traction on land. The tail, instead of flattened for swimming was rounded. due to the center of gravity being well forward of the hips, Pristichampsus was primarily a quadruped, but at higher running speeds could obtain bipedalism.
The oldest known eusuchian is Hylaeochampsa from the Early Cretaceous 130 mya and was already living a modern day crocodilian semiaquatic lifestyle. It is a very primitive eusuchian, but shares a pterygoidal choanae and the absence of a fossa on the medial wall of the supratemporal fenestra as all living susuchians do.

Susisuchus (Phonetics: Su-see-sook-us) is a neosuchian crocodyliform that lies just outside the clade Eusuchia, but is associated with the transition between basal and advanced neosuchians and the rise of early eusuchians.
 |
| Susisuchus anatoceps drawn by: Sergey Krasovskiy |
As recently as the Eocene in the middle of the Paleogene Period (formerly the Tertiary Period) 56-33.9 mya, there was a crocodilian that lived fully on land and ran. Pristichampsus (Phonetics: Pris-stee-champ-sus) had long cursorial legs adapted for running. This was an advantage to being fully terrestrial and on its topside, the jaws were equipped with ziphodont teeth that were characteristic of land predators to grab and tear prey rather than to simply hold onto prey and drown them as aquatic crocodilians do.
 |
| Pristichampsus |
This 3m/9.9ft crocodilian had longer legs than its contemporary semiaquatic crocodilian cousins and in addition, Pristichampsus’s toe ends were hoofed for better traction on land. The tail, instead of flattened for swimming was rounded. due to the center of gravity being well forward of the hips, Pristichampsus was primarily a quadruped, but at higher running speeds could obtain bipedalism.
The oldest known eusuchian is Hylaeochampsa from the Early Cretaceous 130 mya and was already living a modern day crocodilian semiaquatic lifestyle. It is a very primitive eusuchian, but shares a pterygoidal choanae and the absence of a fossa on the medial wall of the supratemporal fenestra as all living susuchians do.
A more derived 100 million year old eusuchian was Isisfordia (Phonetics: I-sis-for-dee-ah) and no it’s not named
after the terrorist group ISIS, but after the nearby town of Isisford,
Queensland, Australia where its fossils were found.
Isisfordia living around 100 mya was a more derived eusuchian than the earlier primitive ones in sharing increased dorsal osteoderm segmentation for more flexibility, ball and socket type vertebrae connections and possessed a secondary palate allowing breathing without passing air through the mouth with all of today’s crocodilians.
 |
| Isisfordia skull |
Isisfordia living around 100 mya was a more derived eusuchian than the earlier primitive ones in sharing increased dorsal osteoderm segmentation for more flexibility, ball and socket type vertebrae connections and possessed a secondary palate allowing breathing without passing air through the mouth with all of today’s crocodilians.
The first true crocodilians were in the
alligator family showing up as Alligatoridae
(Phonetics: Al-la-ga-tor-uh-day) 83 mya. In general, extant alligators have
wider but shorter heads supporting an obtuse snout and the webbing on the hind
feet reach only halfway to the tip. Alligators also cannot tolerate saltwater.
Currently there are two alligator species in the subfamily, Alligatorinae, while Caimaninae is represented by eight extant
caiman.
 |
| Deinosuchus |
 |
| In water, Deinosuchus was a shoe-in to win, on land not so much |
Another extinct caiman, Purussaurus (Phonetics: Pu-ruh-sore-us) lived
as recently as the Miocene 8 mya. Its size was comparable to Deinosuchus with estimates ranging from
10.3m/34ft to 12.5m/42ft. This high-end estimate would make it the largest
crocodylian thus far discovered. Its bite force has been estimated to be 7
metric tons or 15,432 lbs of pressure.
 |
| Purussaurus |
Purussaurus ate large prey by drowning them, as a biochemical analysis indicates it could physically perform the death roll maneuver utilized by the much smaller extant crocodilians of today. In this caiman’s Miocene South American ecosystem, rivers, large floodplains and lake environments existed, offering up many large vertebrates. Unfortunately, the Miocene’s constantly changing environment and climate which began to favor smaller size is a major factor to Purussaurus’ extinction.
One more extinct caiman, Mourasuchus (Phonetics: Moo-ra-sook-us),
was also from the Miocene 8 mya, so was contemporaneous to Purussaurus and also shared the same environment in the Miocene
South American ecosystem. However, Mourasuchus
did not compete with large predators as it was a filter feeder. Like its distant
cousin, Stomatosuchus, this caiman’s
weak lower jaw supported a gular pelican pouch that when it swept or swiped its
open mouth through shoals of small fish or krill, throat muscles would close
and force water through rows of conical teeth on each side of the
jaws acting as a strainer leaving the caiman with a mouthful of food.
The first true crocodiles under the family, Crocodilidae (Phonetics: Craw-cro-dill-uh-day) appeared 55 mya in the Eocene. Crocodilidae is further divided into four subfamilies being, Crocodylinae (crocodiles) with 16 extant species in the genus, Crocodylus, one extant species in the genus, Mecistops; Gavialinae with one extant species in the genus Gavialis; one extant Tomistoma genus species in the subfamily, Tomistominae and two species in the genus, Osteolaemus representing the subfamily, Osteolaeminae.
 |
| Mourasuchus |
The first true crocodiles under the family, Crocodilidae (Phonetics: Craw-cro-dill-uh-day) appeared 55 mya in the Eocene. Crocodilidae is further divided into four subfamilies being, Crocodylinae (crocodiles) with 16 extant species in the genus, Crocodylus, one extant species in the genus, Mecistops; Gavialinae with one extant species in the genus Gavialis; one extant Tomistoma genus species in the subfamily, Tomistominae and two species in the genus, Osteolaemus representing the subfamily, Osteolaeminae.
There are 10 extinct species in Crocodylinae while in the subfamily, Gryposuchinae, all species are extinct. All living Crocodylus crocodiles originated from a
common ancestor from the Indo-Pacific somewhere
~16 mya, radiated out into Africa between 8-12 mya and invaded the
Americas via a trans-Atlantic migration by 4 mya.
Modern day crocodiles can tolerate
saltwater due to the possession of salt excretion glands. Although all feet
have webbing the limbs are tucked to the side using only the tail for
propulsion when swimming. A distinction between crocodiles and alligators are
the narrower snouts forming more of a ‘V’ shape as opposed to a more ‘U’ shape
in alligators.
Although true extant crocodiles have a direct common ancestor going back only 16 mya, the saltwater crocodile, Crocodylus porosus has a direct link to crocodilians going back as long ago as 98 mya when Gondwana had sutured the current Southern Hemisphere continents, which includes Australia, Africa, Madagascar, South America, Antarctica and the India Subcontinent and the Arabian Peninsula with both currently in the Northern Hemisphere. The earliest fossil dates for C. porosus is around 4.25 mya.
Males rarely exceed 6m/19.7ft where the much smaller female seldom surpasses 3.1m/10ft. The record though was pulled from the Bunawan Creek in the Philippines’ province of Agusan del Sur. It measured at a record of 6.2m/20.3ft. C. porosus is not only the largest crocodilian living today; it is the largest extant reptile.
 |
| a) alligator b) crocodile c) gavial |
Although true extant crocodiles have a direct common ancestor going back only 16 mya, the saltwater crocodile, Crocodylus porosus has a direct link to crocodilians going back as long ago as 98 mya when Gondwana had sutured the current Southern Hemisphere continents, which includes Australia, Africa, Madagascar, South America, Antarctica and the India Subcontinent and the Arabian Peninsula with both currently in the Northern Hemisphere. The earliest fossil dates for C. porosus is around 4.25 mya.
Males rarely exceed 6m/19.7ft where the much smaller female seldom surpasses 3.1m/10ft. The record though was pulled from the Bunawan Creek in the Philippines’ province of Agusan del Sur. It measured at a record of 6.2m/20.3ft. C. porosus is not only the largest crocodilian living today; it is the largest extant reptile.
Extant gharials, or gavials and the false gharial have a common ancestor from the Eocene 55mya. These narrow snouted crocodylids are piscivores swinging their jaws back and forth underwater while searching for fish. Once a fish is spotted they quickly snatch it with their sharp interdigitated teeth.

Gryposuchus (Phonetics: Grip-o-sook-us) was a gavial that lived in the Miocene 13.65-7.25 mya beside Purussaurus and Mourasuchus. However, it did not compete directly, as it didn’t go after the larger prey Purussaurus did and did not filter feed small prey as Mourasuchus did; it preyed on fish primarily that were in between the preferred prey size of the other two crocodilians. Also, it grew to comparable lengths at 10m/32.8ft. This gavial was active in riverine and coastal shorelines.

In the next ‘Et Tunc’ series we’ll review the further evolvement of archosaurs within the pterodactyl, dinosaur and bird lines.
Evolving
Onwards,
BJA
01/31/2017
This comment has been removed by a blog administrator.
ReplyDeletepusulabet
ReplyDeletesex hattı
https://izmirkizlari.com
rulet siteleri
rexbet
G72