Et tunc nulla erat VIII
(And Once There
Was)
Dinos: Part 1of 3
The
Path to Dinosaurs:
To set the stage of the environment
dinosaurs evolved from, lived and reigned in, we’ll first start off with period
events that took place during dinosaurs and birds. Below is a description of
the fauna, flora, geological, climatic and catastrophic events during which
time dinosaurs would later evolve as a result of.
 |
| The Early Triassic |
The Triassic is sandwiched in between
two mass extinctions. The first is called ‘The Great Dying’ occurring between
the Permian/Triassic border ~ 252 mya during Pangaea (Pangea). The second
occurred at the Triassic/Jurassic during the initial breakup of Pangaea ~ 201.3
mya.
It’s called ‘The Great Dying’ for a
valid reason...it wiped out 96% of all marine species, 70% of all terrestrial
vertebrates and nine out of every ten plants, destroying Permian conifer
forests en masse. It even exterminated insect groups in killing off 57% of all
insect families and 83% of all insect genera, which included Earth’s largest insects
like the 70cm/27.6in wingspan dragonfly, Meganeura
and other arthropods including the largest land invertebrate ever in the
millipede, Arthropleura. This is thus
far the only mass extinction of insects.
The only surviving vascular land plants
to support any animal life were lycophytes, one surviving ginkgophyta species,
ferns, the seed fern Glossopteris,
some conifers in highlands and bennettitales. However, this opened a window for
spermatophytes (seed plants) to take hold and later dominate in the Cretaceous.
‘The Great Dying’ event was so
catastrophic that from oceanic molluscs to dominant land vertebrates, 96% of
eukaryotic life became extinct. Within 100,000 years, the blink of an eye in
geological time, life was nearly eradicated. There was no one culprit which is
why it has been hard to pin down. But recent paleo stratigraphy analyses have
unearthed the mystery surrounding this mass extinction.
 |
| Falkland Islands Impact Site |
Around 252 mya, an asteroid smashed into
Earth, with remnants of the massive impact crater located near the Falkland
Islands. The crater, now buried in sediment, was first discovered through very
erratic NASA gravity anomalies. Later, magnetic imaging and seismic data helped
verify the 255km/155mi wide crater.
 |
| Falkland Islands Gravity Survey |
This impact was much larger than the 180 km/112 mi
wide Chicxulub impact crater off the coast of the Yucatan Peninsula that finished
off the dinosaurs 186 million years later. ‘The Great Dying’ hyper velocity
impact sent billions of US tons/kilograms (1 US ton = 907.2 kg, 1 billion US
tons = 907,184,740,000 kg) of particle debris into the atmosphere. Along with
the particulates, clouds of noxious gases rose blocking out sunlight for months
on end.
The initial impact on life was exacerbated even more due to another impact around the same period (254.7 ± 2.5). A smaller asteroid, known as the Araguainha Crater slammed into oil shale strata of present day Brazil. In conjunction, these two asteroid impacts shook the globe in tremors unlocking and spewing out escaping underground oil and gas creating massive fires and explosions. Immense coal beds from the Carboniferous were also ignited.
The initial impact on life was exacerbated even more due to another impact around the same period (254.7 ± 2.5). A smaller asteroid, known as the Araguainha Crater slammed into oil shale strata of present day Brazil. In conjunction, these two asteroid impacts shook the globe in tremors unlocking and spewing out escaping underground oil and gas creating massive fires and explosions. Immense coal beds from the Carboniferous were also ignited.
On top of all this, the active ‘Siberian
Traps’ volcanism was jolted into high gear from the asteroid impact. These traps
spewed out basaltic lava covering roughly 2 million square km/770,000 square miles
before its mantle plume played out in the Triassic. Superseding the volcanic ‘Emeishan
Traps’ of present day China that played out at the ending of the Permian 259
mya, the massive volcanism also emitted ash dust, pyroclastic debris and the
gases: hydrogen chloride, hydrogen fluoride, hydrogen sulfide and sulfur
dioxide. Once in the atmosphere, these gases aerosolized in reacting with other
atmospheric components.
This was in effect two pulses of extinction
activity, for of course all the volcanic emissions aided the asteroids’ impact
dust particulates in blocking out sunlight and heat causing a cooling down
effect. The aerosolized gases displaced normal atmospheric conditions turning it
into noxious and corrosive acids that reigned down on Earth as rain and snow.
With sunlight and warmth blocked by the
asteroid impacts and volcanism, the resultant cold darkness and acidification
wiped out photosynthetic plants such as purple algae, green algae and
phytoplankton. These were the base and foundational building blocks of the food
chain. Thus herbivores would starve, which in turn starved out carnivores.
All of this even created a third pulse. The
gases reacting to atmospheric elements such as water molecules, nitrogen and
oxygen produced hydrochloric acid, nitric acid and sulfuric acid that in turn
acidified rain and snow. But the main gas culprit to come was carbon dioxide.
The Permian, although under the ending
of the significant Permo-Carboniferous glaciation, was trending towards a hot
climate during the Late Permian. This was compounded and accelerated in the
Early Triassic with the release of carbon dioxide from the asteroid impacts and
volcanism in the volcanic activity’s initial release of subterranean heat trapped
gases and the releasing of heat trapping gases from the lava flows igniting gaseous
and liquid hydrocarbons that were once trapped in underground rock strata, or
in solid form such as coal veins.

Once all the initial debris and gases were
washed out of the atmosphere, the vast amount of carbon dioxide and carbon
monoxide remained heating up the atmosphere even more. With the abundance of
heat trapping gases in not allowing reflected infrared heat to escape, much of
the excess gases were absorbed by ocean waters. This acidified the waters by
dissolving the inorganic carbon dioxide and carbonates in situ into carbonic
acid deep in the waters due to the fact warmer water sinks. This in turn freed
up sea floor methane hydrate, frozen from the cold and pressure, releasing megatons/mega-kilograms
of methane gas that is 27 times more efficient in trapping heat than carbon
dioxide is. As the Late Permian was already heating up, this accelerated the
process.
Due to this, by the time the Triassic Period was initially underway, atmospheric and oceanic runaway heat was devastating life. Marine species such as blastoids, eurypterids and trilobites, among others all went extinct.
Due to this, by the time the Triassic Period was initially underway, atmospheric and oceanic runaway heat was devastating life. Marine species such as blastoids, eurypterids and trilobites, among others all went extinct.
The more Earth’s climate heated a deadly
march down from glaciation to hothouse incurred. In less than two million years
into the Triassic, there was no evidence of polar glaciers. Even the permafrost
melted with its immense amount of frozen organic debris. Once unfrozen it began
to decompose releasing locked up carbon dioxide. The decomposers were bacteria
and in the process, the bacterium produced methane as waste, further acerbating
the broiling heat up.
How do we know of this...the history of
the episode is literally written in stone and we’ve finally learned how to read
it. We now know this for a few reasons that correlate the time period’s demise
for life. One was in finding and dating the asteroid craters and analyzing
their intensity. In judging the expansive perimeter and mantle depth of the
Falkland Islands impact, its energy force was equivalent to 120.4 billion
Hiroshima A-bombs. This immediately wiped out surrounding life.
Two, after the Falkland Islands impact
intensified the Siberian Traps, the immediate surroundings supporting plants
were wiped out. In addition, the gases and particulate debris added to the
impacts’ blocking out of sunlight, killing off much of Earth’s photosynthetic
plants, thus starving off herbivores, then lastly the carnivores. This is
detected in the increase of fungi spores and microorganisms embedded in the
fossil record that fed off dead organic material. Also, the nickel eating archaeon,
Methanosarcina experienced population
explosions resulting in blooming extensively throughout the oceans.
This archaeon microbe, which still lives
inhabiting the deep oceans and mammal guts, combines carbon dioxide with
hydrogen gas, metabolizing methanol or methylamines producing most of the
biologically-generated methane released today. The nickel content came from the
volcanism that the archaea consumed and metabolized. The by-product of this
metabolism was the release of immense amounts of methane that displaced oxygen
in the oceans while the amount of methane that escaped into the atmosphere
further added to the earth’s trapped heat.

Three, as stated earlier, after all the
debris and gases were washed out of or dissipated from the atmosphere, a vast
amount of greenhouse gases remained heating up Earth’s atmosphere and oceans,
causing a shift in the oceans’ currents. This, in conjunction with the greenhouse
effect, was the long term culprit that killed off life that had managed to
survive through the initial two stages.
Oceans absorb heat energy from the sun
and carbon dioxide from the atmosphere. Oceanic stratified currents move the
energy from areas of hot water to areas of cold water, and vice versa. Not only
does oceanic current circulation keep the water temperature moderate, but it
also affects the temperature of the air and disperses nutrient rich waters in
ocean regions that are depleted.
 |
| Oceanic Current Stratification |
The oceanic current stratification
remains separate due to halocline (salinity), thermocline (temperature), oxygenation
(chemocline) and pycnocline (density) variables. Baroclinity (baroclinic stratified
fluid as measured between the differences of gradient pressure from the
gradient of density) is what maintains a parallel stratification. Wherever they
pop up, upwelling or downwelling eddies create turbulence mixing the stratified
waters. Mixing layers interchange nutrients and acts as a station pump for
current circulation.

When Earth’s water bodies had heated up
due to the runaway greenhouse effects, it began homogenizing the waters’
temperatures, Warmed water rises as cooler waters sink. This is the initial
force in circulating ocean currents. With waters approaching the same
temperature range from the increasing heat, circulation stopped. In so doing,
except along a few coastal shorelines, most of the Panthalassan Ocean and
Tethys Sea bodies became devoid of nutrients and thus void of higher lifeforms.
This created a global dead zone.

First off, when circulation ceased, the
deeper waters became stagnant by losing dissolved oxygen (DO) as warmer waters
released dissolved oxygen content replacing it with carbon dioxide. This
created hypercapnia conditions putting excessive carbon dioxide into the
bloodstream of aquatic animals that lead to breathing problems. This further
led to hypoxic environments, where with little oxygen left animals could not
maintain breathing thresholds so those that could move out did so to shallower
environs. As carbon dioxide further increased, the warming ocean depths further
heated up creating an anoxic environment virtually empty of dissolved oxygen.
For those aquatic animals that were more sessile than mobile they couldn’t swim
away. Unfortunately for marine forms like molluscs, echinoderms and sponges,
they simply perished. In addition, anaerobic bacteria began proliferating in
the depleted DO waters producing toxins that were lethal to aerobic organisms.
 |
| Early Triassic Dead Zone |
However, even where marine animals were
moving into shallower waters just to breathe, most coastal shores were heating
up as well and with the acidification of the waters, corals began dissolving creating
a supported communities die off. Reef making animals during the Permian that
went extinct amid ‘The Great Dying’ were: calcisponge (sponges with calcareous
spicules), algae/bryozoan reef builders and rugose colonial corals. Along the coastal
Tethys seas’ shorelines were about the only escape for marine animals from the
anoxic and heated/acidifying waters.

Almost in a page by page fashion in
flipping through a history book, we know this occurred by observing
fingerprints in the stratified layers of rock and fossils. In an older layer of
bedrock there are many examples of fossils that represent animal deaths, while
the latter formed upper layers of rock show no evidence of fossils.
Except for the terrain higher
elevations, anywhere else on land all the conifer forests perished with a big
influx in fungal spore fossils that decomposed the dead trees. The mean oceanic
temperature along the equator’s perimeter reached a maintained 40°C/104°F.
Terrestrial temperatures ranged from 49°-60°C/120°-140°F. Anoxic conditions from the entire ocean depths
to the interior of Pangaea laid waste in creating a desolate watery grave and desolate
hot desert. Only 4% of all ocean life and 30% of vertebrate terrestrial life
survived.
δ13C did not recover into positive
integers until 244 mya. Life did not rebound until the Middle Triassic and took
thirty million years into the Late Triassic to build up biodiverse ecosystems
and complex food webs.

Isotopic signatures are a valid way in
interpreting, fossils, paleoclimatology, paleoceanography, and geochemistry. δ13C (Delta 13C isotope) and δ18O (Delta
18O isotope) are stable isotopes that occur in nature. The atomic
mass of isotopic variance affects an isotope’s chemical kinetic behavior which
leads to natural isotope separation processes. This is stored in the bedrocks
of former paleo-geographic conditions. Through isotope ratio mass spectrometry,
it tells us what isotopes were laid down in the rock strata at the time, which
in turn tells us what those conditions were.

Above and to the left here, I showed the formulations of the
isotopes, 13C and 18O as ratioed between unstable
isotopes of 12C and 16O respectively. Events like rising
temperatures or biogenic methane isotopes affect the ratios. I’m not going to get
too involved in this as its details could be treated in a long-winded treatise of
its own and I’m sure most of us want to get to dinosaurs. However, I will
attempt to explain a bit of it.
The referenced formulas are the
standards for established material that depict an increase or decrease in the
abundance of the two isotopes and are displayed in parts per thousand (PPT or
per mil); that is what the 1000 ‰ stands for. It will have a negative value when
an unstable isotope supersedes 13C or 18O and will have a
positive value if these two elemental isotopes possess a higher number once
ratioed.
For instance, carbon isotopes are
utilized by plants differentially during photosynthesis, yielding a positive δ13C
value. With this information, we know there was enough plant life to sustain
animal life. But if they yield a negative value, such as large amounts of
methane affecting the mass ratio of 13C to an impinged higher value
to 12C, then we know there was an impact occurring on life. δ13C varies
in time as a function of productivity, C2 abundance, organic carbon
burial, methane amounts and vegetation type.
The same results occur with δ18O. This
oxygen isotope’s negative or positive time shell variances detects atmospheric
and ocean temperatures, evaporation/rain rates, O2 atmospheric/oceanic
volumes and ice sheet transgressions or recessions.
Also, cosmogenic isotopes formed from
the interactions of high-energy cosmic rays and meteorite impacts affect Delta
isotope ratios.
Correlating this information all
together with other isotopic analyses and with the fossil record, a design of
events comes into play. Mass extinctions are signatured by negative δ13C and δ18O
anomalies representing a decrease in primary plant productivity (O2
intake in life) and release of plant-based carbon (sequestered C2
liberation upon death). These laid down isotope signatures embedded in rocks
work just as well for today’s analyses as they do for earlier periods in
Earth’s history and that is telling of what is occurring right now with climate
change.
Study the illustration below. It’s an
interesting comparison of Earth’s atmospheric conditions from the outbreak of ‘The
Great Dying’ to now. In particular compare the earth to 250 mya to the 2090’s.
They are very similar and please recall that ‘The Great Dying’ killed off 90%
of all species. Earth’s abbreviations in the stages are:

P/T ~ The Permian Triassic boundary
LPTM ~ Last Paleocene Thermal Maximum
LGM ~ Last Glacial Maximum
LIA ~ Little Ice Age
A2 ~ Additional Emissions
(Anthropogenic/Manmade)

The above graph, measured in
Milankovitch cycles, is data pulled from Vostok Antarctica ice cores going back
from the present to 420,000 years ago. Milankovitch cycles take into account
and factors in Earth’s cyclical orbit of eccentricity, axial tilt, and
precession. The blue lines on the chart measures CO2 levels, the red
~ temperature range, the green ~ measures methane levels while the bottom orange
line measures δ18O anomalies.
The graph below is a 5.3 million year
change in climate in comparing δ18O oceanic benthic carbonate anomalies to the
Vostok ice cores. Please note between 5.3-3.1 mya there was a warm period due
to high levels of carbon dioxide with a mean global surface temperature
increase of 2-3°C/3.6-5.4°F.

In going back to the last graph, overall
the last three million years experienced cyclic glacial and interglacial
periods. Before that, was a period when atmospheric CO2 levels
ranged from 350-400 ppm heating global climate. Today, as of March 2017,
Earth’s atmosphere is at 406.99 ppm saturation levels, in which by total mass,
is over 850 gigatons. Currently we’re averaging 3 ppm per year increases. At
that rate and with its current exponential gains, CO2 atmospheric
saturation will exceed 500 ppm by 2047. Once this happens global temperatures
will rise 3°C/5.4°F.
What will our world look like at this
increase? Well mankind will still survive, but at an egregious cost. Crop and
food production will be cut short. Due to regions becoming very arid, there
will be massive immigration influxes. There will be a massive uptick in people
dying from heat stroke and regional fresh water supplies will be drained from
extended droughts.
Currently there are some 123.3 million
Americans living along U.S. coastal shorelines. There will be no stopping of
the Greenland and West Antarctica ice sheets from totally melting adding all
that once frozen water into the oceans raising coastal shorelines 10.1m/33ft. With
a mass volume of 0.11 trillion cubic m/2.1trillion cubic ft. of water, compare
that to Mt. Everest weighing 161.9 trillion kg/357 trillion lb. In converting volume
to weight, a NASA and University of California at Irvine analysis already shows
western Antarctica’s current lost frozen water has been equivalent to the
weight of Mt. Everest every two years for the past 21 years. One thing is for
sure the coastal waters are coming.

During the PETM (Paleocene Eocene
Thermal Maximum) ~ 55.5 mya, a large decrease in 13C/12C ratios
of marine/terrestrial carbonates and organic carbon occurred inducing global
warming. CO2 atmospheric and oceanic increases were due to an
averaged atmospheric temperature rise of 6.75°C/12.15°F; although the exact
cause still perplexes geologists. The PETM is a focal indicator for scientists,
because it represents the best analogy of current global warming events with
massive carbon inputs into the atmosphere and oceans, along with the
accompanying acidification of marine environments. The PETM was a runaway mini
hot event that ended as quickly as it came lasting ~ 200,000 years. But in comparing,
it averaged only 0.37 Gt/yr, where currently under human emissions, 10 Gt/yr
are being released.
Below is a NASA video of CO2 in Earth’s current atmosphere. It’s a tad alarming. In particular when there are those in political leadership that have no proof to their denying claims whatsoever, so resort to bashing climatologists, who’ve done the work and once they announce proof from decades of empirical studies, deniers only throw doubt into the winds. Economic greed and wielding power is the only slanted view climate change deniers can offer.
Below is a NASA video of CO2 in Earth’s current atmosphere. It’s a tad alarming. In particular when there are those in political leadership that have no proof to their denying claims whatsoever, so resort to bashing climatologists, who’ve done the work and once they announce proof from decades of empirical studies, deniers only throw doubt into the winds. Economic greed and wielding power is the only slanted view climate change deniers can offer.
Some deniers partially acknowledge
current global warming, but like to claim it is a natural event that has
happened many times before. But they stop there and don’t proceed on explaining
the resultant mass extinctions that ensued. A baked Earth is a dead Earth for
life as we know it. We only have one home...Mother Earth...if we don’t keep the
front porch swept clean there is nowhere else to move to.
After ‘The Great Dying’, by the time
life had rebounded in the forms of archosaurs and synapsids, another mass
extinction event unfolded 201 mya at the boundary of the Triassic and Jurassic.
Primitive dinosaurs had already evolved during the Middle Triassic, but
nonetheless, in this mass extinction an estimated 76-84% of marine and
terrestrial species went extinct. However, the dinosaur line survived to become
the dominant order during the Jurassic and Cretaceous. Massive emissions of
carbon dioxide, sulfur dioxides and aerosols from intense volcanic eruptions
initially created a cooling effect, afterwards an extreme warming period ensued.
This is the consensus in the main energetic force that created the extinction.
The breaking up of Pangaea’s tectonic plate movements created the volcanism
that in turn made up a global environment of both, a critical CO2
greenhouse and a marine biocalcification crisis.

The Cretaceous/Paleogene (formerly
Tertiary) mass extinction unfolded 66 mya wiping out all non-avian dinosaurs.
An estimated average of 76% of all marine and terrestrial species became
extinct. The main significance of the mass extinction was due to the Chicxulub
impactor that will be discussed in fuller detail later on.
 |
| The Chicxulub Crater Asteroid Impact |

As dire as the ‘The Great Dying’ was to Earth’s life, its aftermath on climate change is what promoted the dinosaur lineage evolvement. With only pockets of regional woods left, from once vast interconnected forests and tropical jungles, it isolated species members allowing natural selection to take effect on the surviving archosaurs to evolve biological phylogenetics to better manage the stresses of newer environmental conditions. The further global isolation caused by continental drift then exploded dinosaurian speciation allowing them to enjoy a 180 million year run. Now that is species genetic transfer success...
The Road to Dinosaurs:
The primary tetrapodal survivors of ‘The
Great Dying’ were the synapsids, in particular in the order, Therapsida (Phonetics:
Thur-app-see-duh). In fact, accounting for as many as 95% of the total
individuals in fossil beds, Lystrosaurus
(Liss-tro-sore-us), an herbivore therapsid was the most common terrestrial
vertebrate in the Early Triassic. With a horny beak and the ability to dig for
roots and burrow, gave Lystrosaurus an
advantage in eating varied rough, gnarly vegetation and refuge from the extreme
climate. Also later in the Triassic, bees evolved from wasps in conjunction
with the spread of angiosperms (flowering plants) as insect amber fossils give
evidence to transitional hymenopterans.
 |
| Artist: Julio Lacerda the Therapsid, Lystrosaurus |
Some archosaurian lines managed to
survive as well that eventually led to crocodilians. Poposaurus (Pop-o-sore-us), Prestosuchus
(Press-toe-sue-cuss) and rauisuchids were, among others, sister groups to
crocodylomorphs under the clade, Loricata
(Lore-ee-caw-tuh). Some of the early dinosaurs were on their menu, but it was
the lystrosaurs that sustained dinosaurs during the Early Triassic. They all,
except for the crocodylomorphs went extinct at the end of the Triassic.
 |
| Artist DanOs the archosaur, Prestosuchus |
Another archosaurian line, the
avemetatarsalians also managed to escape extinction during ‘The Great Dying’.
The basal avemetatarsalian, Sceleromochlus
(Sclair-o-mo-klus) was a small insectivore that its ancestral line also
survived the mass extinction. Showing up in the Middle Triassic it lived a
desert life with anatomical features to accommodate a sandy and hot lifestyle
that would be an advantaged inherited pickup for theropod dinosaurs.
 |
| Artist: Mark Witton Scleromochlus |
Sceleromochlus, with a
narrowing of the metatarsal, elongated distal hind limbs and a reduction of the
lateral pedal digits, this creature was a bipedal cursorial animal geared for
speed. For energy conservation in a sandy environ, it was most likely
saltatorial (hopping) as well in locomotion. At the end of the femur was a
pronounced intercondylar groove that supported a large quadriceps femoris
tendon for hopping. At only 18.1cm/7.1in, this avemetatarsalian had a long tibia
and fibula between the knee and ankle and with posteriorly flattened
metatarsals was able to squat or crouch down on the ground in plantigrade
fashion without the feet sinking into sand.
To escape daily heat, it was likely
nocturnal, as betrayed by the very large skull orbits, seeking protective
shelter from the heat during the day. Sceleromochlus
most likely, due to its desert environmental evolvement, possessed filaments or
fuzz, an anatomy trait that pterosaurs and theropod dinosaurs would inherit in
pycnofibers and feathers. Right at the edge of transitioning, Sceleromochlus is a direct ancestor to
ornithodirans.
Below is a Triassic Period graphic of
the animal evolvement lines we’re about to discuss showing when and where they
first evolved.

After archosaurs split into Pseudosuchia (the crocodile line) and Avemetatarsalia (bird-like line) around
249 mya, by 245 mya, avemetatarsalians further divided into two more groups in
the clades, Ornithodira
(Or-nith-o-deer-ah) and Aphanosauria (Ah-fan-o-sar-ee-ah)
evolved. Aphanosaurs are the older of the two clades showing up 245 mya in the
genera, Yarasuchus (Yar-ah-sue-shis)
and Dongusuchus (Don-goo-such-is). The
recent discovery and 2017 description of, Teleocrater
(Tell-ee-awk-rah-tur) also was found in sediment laid down ~ 245 mya. Between
1.8-m/6-10ft long resembling a modern day monitor lizard, it possessed
avemetatarsalian skull features in jaw muscle attachments, but also had
pseudosuchian traits in ankle joints enabling side to side rotation.
 |
| Artist: Mark Witton Teleocrater |
 |
| Simple Ornithodira Cladogram |
Ornithodirans, which includes
dinosauromorphs and pterosauromorphs, had on the fourth trochanter of the femur
an enlarged ridge located low down the ankle joint. This modified and advanced the
metatarsal ankles into a simplified hinge-like arrangement with the astragalus
and calcaneum reduced but firmly attached to tibia and fibula hind leg bones.
 |
| Ankle comparisons |
 |
| Ornithodiran ankles |
Autopodium (plural: autopodia) in
paleontology is the distal endpoint (foot, hands) and measurement of a limb. As
most fossils do not have all four limbs intact, fossil foot imprints left by
the animal is a good way in evaluating, stance, stride, morphology, anatomy, correlation
and identification of species. There are basically two autopodia observed in
fossil studies and that is the ‘manus’ involving locomotion with the forelimb
and an evolving flexing wrist and the ‘pes’ which refers to the hind limbs
including elongation and implanting of the digits.
 |
| A) Manus B) Pes C) Wrist |
Ornithodirans were evolving a middle II,
III and IV tridactyl (three toes) pes elongation. Eventually, even quadruped
sauropod and ornithischian dinosaurs would show a pes elongation with
lengthened and symmetrical tetradactyl (four toes) plantigrade pes autopodium
form, along with an asymmetrical tridactyl manus form. Birds go all the way
back as the only surviving ornithodiran group.
 |
| Pes three-toed elongation |
 |
| Marasuchus skeletal |
 |
| Artist: KirbyniferousRegret1 Marasuchus |
Marasuchus’ digitigrade
bipedalism relied on a tridactyl pas elongation, even though it could move as a
quadruped. This animal may also have had filamentous integuments or even
proto-feathers of simple shafts which matted together in covering the body.
This would suggest an insulating function, further suggesting endothermy. Both,
from primitive to the most diverse pterosaurs and dinosaurs inherited these
traits as we’ll soon comment on for dinosaurs later on.
Dinosauromorpha:
Dinosauromorphs were more closely
related to dinosaurs than to pterosaurs, but were not dinosaurs. However, they
gave rise to dinosauriforms, which in turn gave rise to dinosaurs. Basic
dinosauromorph osteology is that the femur head has very little offset from the
shaft, in particular when compared to dinosaurs. They stood erect and moved in
parasagittal fashion in swinging the limbs fore and aft as opposed to rotating them.
Dinosauromorph shoulder assemblies were robust with simple and triangular hip
girdles.

Dinosauromorphs showed up in the Middle
Triassic ~246 mya, but in high latitudes, lived with dinosaur faunas throughout
the Late Triassic for 20 million years disproving any rapid ecological
displacement between the two groups.
The 235-211.9 mya dinosauromorph family,
Lagerpetidae (La-grr-peat-a-day) was
composed of three genera being, Lagerpeton
(La-grr-pee-tun), Dromomeron (Druh-mom-rin),
and Ixalerpeton (Ik-sal-ur-pee-tun). Lagerpetids
were a sister group to dinosauriforms.


 |
| Artist: Apsaravis Lagerpeton |
This family group is defined by having posterior
dorsal vertebrae with anteriorly-oriented neural spines, a sacrum with two
vertebrae united to the ilium with a well-defined anterior projection, a wide
pubis shorter than the ischium and laminar, sigmoid femur with pneumatization
and a well-developed fourth trochanter without phalanges, metatarsal I short
and V reduced, two distal tarsals corresponding to metatarsals III and IV with
metatarsal IV longer than the rest. The acetabulum had closed with an up facing
ilium; traits that would carry into dinosaur groups.
Dinosauriformes:
Dinosauriformes encompasses the
dinosauriform as the common ancestor to all dinosaurs and dinosaurs themselves.
Dinosauriforms all share shortened forelimbs, three or more sacral vertebrae, a
partially to fully perforated acetabulum, an extension of femoral articular
surface under the head of the femur, a cnemial (tibia/shinbone) crest on
proximal tibia with the tibia also being posteriorly flanged on its distal end.
Dinosauriforms that weren’t quite a
dinosaur are mostly under the family, Silesauridae
(Sile-o-soar-uh-day) with the outside genus, Marasuchus (Phonetics: Mah-rah-su-cuss) as representing a sister common
ancestor to the group of silesaurids. Thus far, Marasuchus is monophyletic representing no family group and has
only one species in, M. lilloensis
(Phonetics: M. lill-o-in-sis).
Marasuchus had no acetabulum (hip socket hole) as true dinosaurs did, but it did share an elongate pubis and an anterior trochanter on the femur. Since Marasuchus lived 235-234 mya, dinosaur evolving traits showed up during the Middle Triassic. Lightly built and small at 30-40cm/12-16in, Marasuchus was also a bipedal predator preying on small vertebrates and large invertebrates. In paving the evolutionary route, the earliest dinosaurs were also lightly built and traversed bipedally.
Marasuchus had no acetabulum (hip socket hole) as true dinosaurs did, but it did share an elongate pubis and an anterior trochanter on the femur. Since Marasuchus lived 235-234 mya, dinosaur evolving traits showed up during the Middle Triassic. Lightly built and small at 30-40cm/12-16in, Marasuchus was also a bipedal predator preying on small vertebrates and large invertebrates. In paving the evolutionary route, the earliest dinosaurs were also lightly built and traversed bipedally.
Since at least some early dinosaurs had
primitive feathers (proto-feathers), often times Marasuchus is pictured as feathered, although there as yet has not
been any definitive fossilized feathering imprints found.

The Silesauridae
family, as a sister group to dinosaurians, currently consists of six genera. They are: Lewisuchus, Asilisaurus, Eucoelophysis,
Silesaurus, Sacisaurus, and Diodorus (Phonetics:
Lew-iss-su-cuss, A-se-lee-sore-us, Yew-see-lo-fy-sis, Sy-el-sore-us, Sa-suh-sore-us,
Dee-uh-dor-us). Pseudolagosuchus
fossil remains are fragmentary and thought is trending that it is now synonymous
to Lewisuchus.
Silesaurids were all relatively small and basically walked in a plantigrade quadruped fashion, but could rear and run bipedally if needed. Silesaurids commonly possessed a brevis shelf (muscle attachment site on ilium bone surface), a slender shafted ischium and a prominent lesser trochanter, which were all dinosaurian characteristics as well.
Although derived from carnivores, most
silesaurids were herbivores, likely due to a conferred evolutionary advantage
in what was going on during this portion of the Triassic when all the
continents were still conjoined and Earth and life were still recovering from
the mass extinction. The shift in diet of these dinosauriform ancestors to more
readily available foods led to an eventual dinosaurian success.
As a family clade, silesaurids lived
between 245-203 mya in the Middle and Late Triassic. Appearing 245 mya, Asilisaurus is one of the oldest known
animals of the dinosaur/pterosaur avemetatarsalians. Silesaurus fossil remains are also from 245 mya, but also occurred
all the way into the end of the Triassic 203 mya, well after true dinosaurs
began walking the earth.
Asilisaurus (245 mya) ~ At 1-3
m/3-10ft, this dinosauriform was a quadruped. By judging its dentition, it was
also an omnivore capable of eating plants mostly, but occasionally meat. Ending
in a beak-like toothless tip, the rest of the jaw teeth were triangular shaped,
while large in front getting progressively smaller down the jawline. With the
remains of Asilisaurus found in a
Tanzanian fossil bed, including primitive crocodilians alludes to the fact that
these two animal groups diversified rapidly into their respective dinosaur and crocodile
lines. Asilisaurus was an herbivore
and is one of the oldest archosaurs leading to the ornithosuchus line of
pterodactyls, dinosaurs and birds.
 |
| Artist: Andrey Atuchin Asilisaurus |
 |
| Artist: Scott Reid Lewisuchus |
Lewisuchus (236-234 mya) ~
At 1m/3ft, it was another quadruped that differed from other dinosauriforms in
dental, vertebral and skull anatomy. Lewisuchus
had pterygoid teeth, postaxial neck/trunk vertebrae with craniocaudally
expanded neural spines and a dorsomedial post-temporal opening on the otoccipital
portion (posterior half of the braincase in basal tetrapods) of the skull. The lower
jawline dentary scheme and forelimb anatomy exhibits autapomorphic
modifications related to predatory behavior. Lewisuchus was carnivorous preying on small vertebrates.
 |
| Artist: Apsaravis Silesaurus |
Silesaurus (230 mya) ~ At 2.3m/7.5ft,
was also a quadruped, but due to the presence of three sacrals firmly connected
to ribs and with the ilia and long tail providing a counterbalance to the body
weight in front of the pelvis, suggests an ability for fast bipedal running. Although
it lacked femoral and dental traits of Dinosauria
members, this dinosauriform had the same characteristics of ornithischian
dinosaurs in possessing the beak tip. Also, as evidenced from its bone
characteristics, had a rapid maturation process, but with dense more
longitudinal vascularization ceasing before reaching the peripheral avascular
parallel fibered bone formed at the bone’s surface. This is an indication of a growth
halt and significant decrease in the maturation ratio. The same observation is
found in fossil research of all ornithischians along with most other dinosaurs.
Silesaurus was herbivorous.
 |
| Artist: Apsaravis Eucoelophysis vs. Redondasaurus |
Eucoelophysis (228-208 mya) ~
At 3m/10ft, was still another quadruped but with much shorter and gracile
forelimbs. Originally it was considered a primitive coelo-physoid dinosaur but
its hind limb anatomy negated that thought. The undeveloped posterior femoral
notch and the tibia possessing a distinct appressed surface along the distal
two-thirds of the bone while lacking a fibular crest, are traits too primitive
to be a dinosaur. It’s now considered a sister taxon to Silesaurus. Eucoelophysis
was a carnivore.
 |
| Artist: Nobu Tamaru Sacisaurus |
Sacisaurus (225 mya) ~ At
1.5m/4.9ft, its osteology was very similar to ornithischian dinosaurs. There is
a process at the tip of the mouth resembling the ornithischian predentary bone.
With teeth no longer than 3mm/0.12in this dinosauriform could not adequately defend
itself with biting teeth. However, the teeth were excellent for processing
plant material. The legs were long in proportion to body length indicating an ability
for speed in running. Sacisaurus was
an herbivore.
 |
| Artist: Scott Reid Diodorus |
Diodorus (216 mya) ~ At
< 1m/3.3ft, it had forward canted teeth that decreased in size toward the
anterior end of the lower jaw. Also with smaller teeth, there was a distinct
lateral ridge running parallel to the dentary alveolar margin. Diodorus is considered as the sister
taxon to Sacisaurus. The Diodorus taxon provides further evidence
of a near-cosmopolitan range for basal dinosauriforms in the Late Triassic and
further demonstrates the disparity of dental morphologies getting smaller within
Silesauridae. Diodorus was herbivorous.
Dinosauriforms had set the stage for
true dinosaur evolvement as their fossil finds throughout the globe are
evidence of a near-cosmopolitan range for dinosaurs to thrive in by taking
advantage of natural selective biomes, ecologies and environments.
Dinosauria:
The first of true, but primitive
dinosaurs to appear were around 243 mya, moving the dinosaur lineage back
another 13 million years than what was previously thought. So we cannot think
that dinosaurs were a homogenous pot occurring as all together during a certain
period. When dinosaurs first appeared, they did not facilitate a rapid takeover
in out-competing other species in hastening their extinction. Dinosaurs lived
side by side among other animal species groups, such as crocodilians,
synapsids, sphenodonts, Rauisuchus,
rhynchosaurs, pterosaurs dinosauromorphs and dinosauriforms.


In fact, from a 230 mya swamp fossil
bed, embedded in its rock, were remains of the basal sauropod dinosaur, Buriolestes alongside the dinosauromorph,
Ixalerpeton and numerous other animal
groups.
 |
| Artist: Maurílio Oliveira |
In the illustration above, it includes: Buriolestes climbing over the fallen
tree trunk with six others behind it, seven Ixalerpeton
in the lower left corner, an aetosaur in the lower right corner, a small
rauisuchian to the right center, a sphenodont on the tree and a yellow
amphibian entering the water in the right-center.
The shared anatomical features of
dinosaurs are:
1)
A
supratemporal fossa (skull excavation).
2)
An
epipophyses present in the anterior neck vertebrae behind atlas and axis.
3)
Radius
(a lower arm bone) 80% shorter than humerus bone length.
4)
Femur's
(thighbone) fourth trochanter is a sharp flange.
5)
4th
trochanter asymmetrical with distal lower margin forming a steeper angle to shaft.
6)
Exoccipitals
(bones at the back of the skull) do not meet along the midline on the floor of
the endocranial cavity as in former archosaurs.
7)
Proximal
articular surfaces of the ischium with the ilium and the pubis are separated by
a large concave surface.
8)
Upper
ankle bones and the proximal articular facet (connecting surface for fibula)
occupy < 30% space of the transverse width of the element.
9)
The
pelvis proximal articular surfaces of the ischium, ilium and pubis are
separated by a large concave surface.
10) Tibia’s
cnemial crest arcs anterolaterally.
11) Distinct vertical oriented ridge on posterior face of tibia’s distal
end.
12) Top surface of the calcaneum where adjoined to fibula has a hollow
profile.
As well, Dinosauria are now considered
monophyletic due to numerous autapomorphies including: elongate vomers, three
or more sacral vertebrae, three or fewer phalanges on fourth digit of manus,
grasping hand produced by a thumb that could bend inward (bending occurs at
joint between metacarpal I and first thumb phalange), fully offset (inturned)
proximal head of femur with a distinct neck and ball, greatly reduced fibula,
well-developed ascending process of astragalus, upright posture and a digitigrade
stance.
Traditionally, dinosaurs have been
divided into two subgroups known as, Ornithischia
(Or-nee-this-key-uh) and, Saurischia
(Sawr-is-key-uh). The two clades’ main dinosaurian differences are in the hip
joints, where ornithischians’ pelvis is bird-like, while the saurischian pelvis
is lizard-like. Ornithischia is Greek
and actually means bird hipped, where, Saurischia
in Greek means lizard hipped. Saurischia
is further divided into two subgroups being, Theropoda and Sauropoda.
This all has to do with the positioning of the pelvic bones, the ilium, ischium
and pubis. Ornithischians’ pelvis is more situated as birds, while saurischians
have the three bones positioned more like lizards. The bird hip is considered
an ‘opisthopubic’ pelvis, where, as in birds, the pubic bone extends back
towards the tail of the animal. A lizard hip is a ‘prepubic’ pelvis where the
pubic bone extends forward towards the animal’s head. This is where I want to
chime in on a thought before we go any further.

Above are the familiar dinosaur hip
illustrations we’re all familiar with. As but a young fella, I’ve always
wondered how theropods, which indeed were lizard hipped saurischians could have
evolved into birds when it was ornithischians that had the bird hipped
arrangement. Well, it appears that finally paleontologists are beginning to
figure this out.

Above is a new cladogram devised by
Ph.D. Baron Matthew when he was a doctorate student at the ‘University of
Cambridge’. In the illustration, are the phylogenetic relationships of early
dinosaurs through a time-calibrated strict consensus of 94 trees from an
analysis with 73 taxa and 457 characters. A – Is the least inclusive Dinosauria clade that includes Passer domesticus, Triceratops horridus and Diplodocus
carnegii. B – Is the least inclusive clade of Ornithoscelida (Phonetics: Or-nee-tha-o-sell-lee-duh) that includes
P. domesticus and T. horridus. C – Is the most inclusive Saurischia clade that contains D. carnegii, but not T. horridus.
The cladogram further illustrates all
subdivisions of the time periods (white and grey bands) are scaled according to
their relative lengths with the exception of the Olenekian (Early Triassic),
which has been expanded relative to the other subdivisions to better show the
resolution within Silesauridae and
among other non-dinosaurian dinosauromorphs.
What this tells us is that as more
dinosaur discoveries are being made, phylogenic and anatomical relationships
have made us realize that the simple hip bone categories are no longer adequate
enough to separate dinosaurian relationships. The traditional
bird-hip/lizard-hip relationship is no longer adequate in the thinking that
ornithischians and saurischians came from separate ancestors. Now we know that
dinosaurs all evolved from a common ancestor.
However, in more ways than one
ornithischians are anatomically more similar to theropods than theropods are to
sauropods. Further, in coming from the same clade, ornithischians and theropods
both had the same potential to develop bird-hip anatomies; it’s just that the
ornithischians did it first, before theropods hit the genetic switch when they
began evolving into birds. This revised clade grouping of ornithischians and
theropods is a revival of the name, Ornithoscelida,
originally coined by evolutionary biologist, Thomas H. Huxley in 1870. In this
scheme, herrerasaurids as a basal sister group to sauropods, are both grouped
under Saurischia. This makes
sauropods the least related dinosaur to ornithischians and theropods. Sauropods
simply retained their ancestral archosaur reptilian pelvic arrangements. Also,
this cladogram firmly attaches theropods as the basal common ancestry to all
birds extant and extinct.
Please keep this in mind, for it may not
yet be consensus, but as the majority of paleontologists have the Baron
cladogram in waiting for more review, soon I feel, it most certainly will be
the new dinosaur clade. Although this Baron et al grouping only uses species
level operational taxonomic units (OTU) that enlists metagenomics in utilizing
similar sequence variants, which in itself has a few limits in distinguishing
at the species level, I will be introducing dinosaur clades under the new Ornithoscelida clade scheme instead of
the older current ones.
 |
| X - Out with the old cladistics and in with the new - ✔ |
The early dinosaurs, as exposed from
fossil findings, had a large variation which most likely allowed higher
successful rates in harsh environmental conditions that the Triassic offered.
One main variation was growth rates within species. From ontogenetic sequence
analyses, early dinosaurs had a much higher level of growth patterns than did
the other reptile groups at the time. In addition to possessing different growth
pathways from hatchling to adult, there too, was a large amount of size
variation within species like a younger adult individual being much bigger than
an older individual. This is the same biological strategies humans possess as
well, for in any community you will find much taller teenagers compared to
grown adults.
As a result, when the Triassic/Jurassic
extinction occurred, the species and individuals of species with the most variants
to withstand the harsh environmental conditions managed to survive and those
that didn’t, succumbed. Except for crocodilians and dinosaurs, all archosaurs
went extinct at the Triassic/Jurassic border. Other groups also perished as
displayed in the extinction graph below.
 |
| Triassic/Jurassic Extinction |
An interesting basal archosauriform that
didn’t make it past the extinction event was Vancleavea (Phonetics: Van-cleve-uh). At 1.2m/3.9ft log, it was
semiaquatic and had a tail deepened by elongated osteoderms as opposed to tall
neural spines that was normal in all other aquatic tetrapod groups. Two other
unusual traits were imbricating osteoderms covering the entire body and the
jaws were lined with enlarged caninform teeth (non-mammalian animals that
possess dog-like canines) that protruded from the mouth.
 |
| Artist: Smokeybjb Vancleavea |
 |
| Artist: Yu Chen Limusaurus |
Limusaurus, a biped stood
at 1.5m/4.9ft when fully grown. Limusaurs came into the world with a full set
of firmly rooted carnivorous teeth, but as it aged it lost all of its teeth as
they were replaced with an herbivorous beak in adulthood. As yearlings, limusaurs
were fully carnivorous going into the subadult stage. At one year it began
losing back teeth foregoing carnivory for an omnivorous diet until reaching
adulthood. As an adult, all teeth were replaced by a beak in favoring an fully
herbivorous diet.
Apparently this kept competition at bay when
it came to food resources between the young and adult limusaurs. To accommodate
the diet transitions, proportions of the skull and upper jawbone configuration
began to modify in a pronounced downward curve to develop the exterior beak.
The larger adult individuals swallowed stones to aid the gizzard in grinding
vegetative food, whereas the carnivorous young did not.
Newer speciation arose from within a
species through vicariant speciation which is allopatric (geographic
speciation). This form of speciation emanates when biological species become
isolated through geographical means. A classic example is in the theropod family
of carcharodontosaurids.
 |
| Artist: Dinoraul Acrocanthrsaurus |
In the Early Cretaceous, Acrocanthosaurus (Ack-row-can-fo-sore-us)
showed up around 116 mya. At that time, the world’s landmasses were still
enough connected allowing for transcontinental dispersal. Acrocanthosaurs originated
in what is now the central southern portion of the U.S. in Texas and Oklahoma.
This dinosaur spread throughout what is now South America and Africa. By the
Late Cretaceous land bridges intra landmass contacts had been taken over by the
oceans and the further continental spreading.
 |
| Artist: Willydynamo55 Giganotosaurus vs. Macrogrypgosaurus |
This isolated acrocanthosaur populations
in varying ecological environs of the forming continents, to the point the
South America population of Argentina developed enough genetic variation to
become the new genus, Giganotosaurus
(Gye-guh-nuh-tah-sore-us) as the result 98-97mya. The same happened in the
North African population forming the genus, Carcharodontosaurus
(Car-kah-roe-don-to sore-us) 112-93.5 mya. The ecological isolation further
affected size, with Arcocanthosaurus
at 11.5m-38ft, Giganotosaurus at
12.5m/41ft and Carcharodontosaurus at
~ 13.3m-44ft.
 |
| Credit:BBC Carcharodontosaurus |
Like pterosaurs, skeletal pneumaticity
was found in dinosaur fossils. Functions of skeletal pneumatics include weight
reduction in large-bodied or flying dinosaur taxa and density reduction
resulting in energetic savings during foraging and locomotion. The pneumatic
system is comprised mostly of the lung. Indeed, all the little pockets that
arise come from expansions of the primary bronchus, a tissue that surrounds the
lung itself. It is this tissue forming the little sacs. This is known as the
‘pneumatic diverticulae’ that ends up throughout the body as well as inside the
bones.

Though evidence is lacking in caudal
pneumaticity in sauropod fossil finds, it is quite evident in the rest of the vertebral
column. Also, in all sauropods, air-filled hollow bones were throughout the
postcranial and anterior body. Like dinosaurs’ nearest surviving relatives of
crocodilians and birds, sauropods did not have a diaphragm, so the dead-space
volume of a 30-ton sauropod would be about 184 liters of air. This is the total
volume of the mouth, trachea and air tubes. If the animal exhales less than
this, stale air is not expelled and is sucked back into the lungs on the
following inhalation. This is how we know that sauropods relied on respiratory
pneumatics to properly inhale and exhale.

Pneumaticity of the cervical and
anterior dorsal vertebrae occurred early in theropod evolution. Although not
pinpointed, evidence shows that only some of the theropod thoracic air sacs
were present as plesiomorphic, but then modified and embellished on by each
theropod clade separately. The acquisition of extensive postcranial
pneumaticity in small-bodied maniraptorans (clade consisting of birds and
non-avian dinosaurs) makes avian-like high performance endothermy a good
sequel. Oviraptorids (bird-like, herbivorous and omnivorous maniraptoran
dinosaurs) showed an astounding number of skeletal air sacs and postcranial
pneumaticity sites.

Flow through ventilation first occurred
in theropods. Due to an incredibly preserved Upper Cretaceous ‘Maevarano
Formation’ fossil find of Majungasaurus
(Muh-jung-ah-sawr-us) in NW Madagascar, the series of vertebrae and portions of
the ribs preserved pneumatic foramina (cavities) that may have resulted from
the infiltration of avian-style lungs and air sacs. This would have been a
basic ‘flow through ventilation’ system where air flow through the lungs is
one-way allowing oxygen-rich air inhaled from outside the body to never be
mixed with exhaled air laden with carbon dioxide. This is the same breathing
system birds of today extol and although it’s complex, it is highly efficient. Majungasaurus roamed 70-66 mya.

Carcharodontosaurus, mentioned
earlier above, utilized ‘flow through ventilation’ to the maximum in efficiency
by employing a row of pneumatic air sacs so devised that it ensured a
continuous and constant flow of oxygen be supplied to and through the lungs
during respiration. This gave this dinosaur a huge advantage over its fastest
prey, as it could easily outrun them. It also complimented its hunting style,
which was viper like to strike and recoil, then wait for the wounded prey to
die. Carcharodontosaurs, with large forelimb claws or a quick bite, would lacerate
or slice its shark-like serrated teeth into a vulnerable area, wait out for the
mortal wound to take effect then begin feeding on the dead prey. This strategy
saved this carnivorous theropod from any injuries that might occur during
battle with a struggling prey that might have had its own weaponry of
defenses.
However, concerning ornithischians, there
has thus far been no evidence of skeletal pneumaticity found in their dense
bones. Perhaps they had developed intricate lungs for air capacity or soft
tissue air sacs that wouldn’t fossilize. Ornithischians show an evolving
reduction in skull pneumaticity with the closing of the antorbital cavity.
Reasons for this could be as the feeding skull components expanded
phylogenetically, the antorbital cavity contracted. Or simply, weaker point
skull pneumaticity gave way as bonier head components and increased body weight
arose for defense.
As far as intelligence goes dinosaurs
were not the lumbering dimwitted nitwits we once thought they were. All
dinosaurs could be quick reacting under stressful situations, even mental reasoning
to cause and effect. One really stands out when it comes to intellect and
judgment. Occurring 77.5-76.4 mya, Troodon
(Tru-o-don) was perhaps the most intelligently derived dinosaur thus far
discovered. Its cerebrum to brain ratio was up to 63% higher than other
non-avian theropods. The brain itself as compared to body size was much larger
than extant reptiles and most probably possessed the same measured intelligence
as most birds.
 |
| Mugger crocodile using sticks as tools for bait |
We don’t know if dinosaurs used simple
tools as today’s American alligator or mugger crocodile does in stacking sticks
on top of its snout to entice then snare breeding shorebirds, but Troodon certainly had the intelligence
to do so.
In diet, most of the earliest dinosaurs
were insectivores and carnivores, with a few omnivorous species and very little
herbivores. Of course once into the Jurassic in kicking off their reign,
dinosaurs covered every base in being insectivores, herbivores, carnivores, scavengers
and even with spinosaurs, became semiaquatic piscivores.
Even cannibalism is evident in, Majungasaurus. Unlike
carcharodontosaurs, majungasaurs with strong jaws and a wide bite filled with
gripping teeth, would hold prey with the bite. With the exacting majungasaur teeth
marks found in the fossils of other majungasaurs, we know it cannibalized, but
we don’t know the reasons why. Prey scarcity due to changing environmental
conditions could’ve made it desperate for food or it could’ve come upon an
already rotting dead majungasaur carcass. Thus far though, Majungasaurus is the only genus of dinosaur found to eat its own
kind.
All carnivorous theropods would tear off
chunks or strips of flesh from their prey and swallow it whole. Primitive ornithomimosaurs
and oviraptorosaurs evolved from carnivorous theropods, but with weak jaws and
small dulled teeth probably were omnivorous.
 |
| Artist: Damir Martin Therizenosaurus herbivorous family |
One group of theropods that did evolve
from a flesh eating carnivore into a plant eating herbivore was in the family, Therizenosauridae (Phonetics:
Thu-rise-zen-o-sawr-ah-day) with species living 94-66 mya. These were large
theropods that reached up to 10m/33ft in length. With very broad bodies and
long necks standing on weight bearing hind limbs supported by four weight
bearing toes, was an unusual evolvement from its contemporary theropods that
walked on three toes with the fourth representing a dew claw.
Even though this has occurred throughout
the course of Earth’s biological history, such as hominids going from a
strictly herbivore diet to an omnivore one, or even a fully carnivore one as
some West Texas cattle ranchers would like to attest to, for a flesh eating theropod
converting over to a plant diet is unusual upon itself in the body
accommodating the transitioned diet.
 |
| Therizenosaur manus claws |
But what made therizenosaurs even more
remarkable from other theropods with normally short forelimbs was in the size
of their three forelimb claws ending on rather long arms. At just under
1m/2.2ft, therizenosaurs had the longest claws of any known animal extinct or
extant. In being considered a top contender for the emergence of an herbivory
lifestyle within Theropoda,
therizenosaurs had short, but packed serrated teeth for slicing and chewing in
a small mouth. With the theropod defense gone in its biting abilities,
therizenosaurs evolved and developed the large claws ending on robust and long
arms for defense, although the longer arms and claws may have originally
evolved to reach for higher limbs, clamp onto them and bring them down towards
the mouth. Just as in venomous snakes, where the toxin from modified salivary
glands was first developed for capturing prey and as a secondary element for
defense, so too might have been the therizenosaur forelimbs.
In jumping ahead here, as this bit of
info should be addressed later on under the bird section, but in considering therizenosaurs’
evolvement into herbivores; due to their theropod anatomy, certain morphologies
evolved that may have started the trend towards theropod birdom. A keratinous
beak tipped rostrum was developed to enhance cranial stability by mitigating
the stresses and strains of chewing in pre-processing vegetation material and
cellulose. For a more adaptable upright reach, the pectoral girdle was modified
to augment longer reaches. Therizenosaurs also evolved an opisthopubic pelvis
(pubic bone extends posteriorly) much like ornithischians and birds. This was
done to enhance the weight bearing hind limbs. But most of all, (although not
for flight, instead, for thermoregulation and courtship), therizenosaurs are
one of the first animals to produce bird-like feathers; albeit primitive feathers
considered as stage-1where the feathering was spine-like. The latter more
evolved therizenosaurs possessed stage-3 serially fused feathering barbs.
Herbivore dinosaurs had jaws that only
slightly opened and closed where all the teeth met to masticate the food,
except for sauropods; they only had teeth suitable for stripping food items
from plants then swallowing it whole relying on large abdomens equipped with
gastroliths (swallowed stones) to grind and bacteria to aid in the digestion of
cellulose. Other herbivores like most ornithopods had no teeth but were
equipped with horny beaks to snip off food items then swallow it whole and
allow the digestive tract similar to sauropods to break down food tissue. With
no large stomachs to accommodate vegetative materials, the beaked ornithopods,
such as hadrosaurs had beaks for snipping and weak jaws and teeth for grasping,
but not for chewing. They most likely ate fruits and softer plant materials not
requiring an elaborate digestion tract.
 |
| Chilesaurus skeleton |
 |
| Chilesaurus jaw and teeth |
 |
| Artist: Gabriel Lio Chilesaurus |
During their reign, large carnivorous
dinosaurs ate pretty much any other animal during their dominance in the
Jurassic and Cretaceous, but any of them could have also been prey by being on
other carnivorous animal’s menu.
 |
| Artist: Fabio Manucci Razandrongobe |
During the Jurassic lived, the oldest
known notosuchian, Razanandrongobe (Ray-zah-non-dron-goab),
a crocodyliform living 167-164 mya. It possessed teeth larger than any predator
dinosaur of the day and were used for crushing bone. It was an apex predator that
could subdue any dinosaur during its time.
 |
| Artist: Sergey Krasovskiy Beelzebufo |
Other animal predators that ate
dinosaurs were crocodylians, pterosaurs and even a particularly large toad
known as, Beelzebufo (Be-l-zah-buff-fo)
could easily have handled many types of baby dinosaurs.
 |
| Artist: Mark Witton Quetzalcoatlus |
 |
| Artist: Mark Witton Hatzegopteryx |
The pterosaur, Hatzegopteryx (Hat-zee-gop-ter-ix) also must’ve dined on dinosaurs. Isolated islands make large species shrink in size and small species enlarge in size. This phenomenon is called the ‘island rule’. During the Cretaceous, there existed small European subtropical islands during the break up of Pangea within the Tethys Sea. These islands had animal populations separated from their mainland kin. The sauropod, Titanosaurus (Tie-Tan-o-sawr-us) was large on the mainland grounds measuring up to 12m/40ft. Isolated from the mainland on one particular island called, Hateg Island that is now a part of Romania, titanosaurs experienced insular dwarfism cutting their size in half to no more than 6m/20ft. Their young weren’t even half that size. In fact the island titanosaurs were so miniature in size as compared to mainland titanosaurs that they’ve been assigned another genus, Magyarosaurus (Mag-yar-o-sawr-us). Hatzegopteryx would soar from the main lands to the island where the magyarosaur young were easy pickings. Hatzegopteryx would also prey on other dinosaurs small enough to catch and swallow in one gulp.
 |
| Artist: Robert Nicholls Deinosuchus |
 |
| Artist: Josh Cotton (greeni-studio) Deinosuchus |
 |
| Artist: James Guerney Repenomamus |
Even mammals of the day took their share of small dinos. Living 125-123.2 mya, Repenomamus it is the largest Mesozoic mammal known at 1m-3.3ft long. It was carnivorous and at one of its fossil finds in the ‘Yixian Formation’ of China were the remains of a young Psittacosaurus in its stomach.
 |
| Repenomamus fossil with Psittacosaurus remains |
Dinosaurs roamed throughout the Jurassic
and Cretaceous world topographies. Fossils have been discovered in lowlands,
highlands, deserts, forests, tropical climes and cold climes in both the Arctic
and Antarctic. During Cretaceous ice free summers, the earth poles experienced
only intermittent sea ice throughout the rest of the year. Average pole temps
were 10 °C/18
°F because
there was much higher concentrations of CO2 in the atmosphere at
1000 ppm…two and a quarter times than currently, even though today’s
atmospheric CO2 saturation levels are alarmingly increasing. Even
though in today’s arctic temperatures, 10 °C/18 °F would be mild weather that
range for a reptile is still cold. Most extant ectothermic reptiles cannot
tolerate higher latitude arctic conditions as their survival is dependent on
ambient temperatures.
Let’s get reacquainted with organism
body regulations:
§
Poikilothermy
~ internal body temperature highly varies
§
Homeothermy
~ maintaining a stable internal body temperature regardless of external
influence
§
Stenothermy
~ survival only within a narrow temperature range
§
Eurythermy
~ can function at a wide range of different body temperatures
§
Ectothermy
~ relying almost purely on ambient heat
§
Endothermy
~ heat set free by internal bodily functions
So for dinosaurs to have existed in cold
environments, they could not have been poikilothermic/stenothermic. Dinosaurs
that inhabited the pole regions were either homeothermic/stenothermic, or
endothermic/eurythermic, or at least be poikilothermic/ectothermic as the mammalian
naked mole rat is today.
 |
| Fossilized Haversian canals |
Also, we now realize dinosaurs weren’t
as cumbersome and sluggish as an ectotherm (cold blooded) would be. Dinosaurs
were very active and agile with higher metabolic rates than an ectotherm would
be. The reasons we now feel dinosaurs were endothermic (warm blooded) is for
one, their bones possessed Haversian canals which are a series of microscopic
tubes containing capillaries and nerve fibers. The bones possessing the canals
are called cortical bone with the Haversian canaling occurring in the outermost
region of the bone. The only animals we know of that also possess Haversian
bone are mammals and birds, in which both are endothermic.

Dinosaurs lived and died near both poles
enduring not only the cold, but endless winter days of no sunlight. A hadrosaur
fossil find of Edmontosaurus (Ed-mon-toe-sawr-us)
was discovered near the North Pole. In other fossil finds, there was the
discovery of a new hadrosaur species, Ugrunaalluk
kuukpikensis, meaning ‘ancient grazer in Inupiat Eskimo. With other
hadrosaurs, pachycephalosaurs and thescelosaurs fossils, there were also the
carnivorous dromaeosaurs, gorgosaurs and troodontids. The Alaskan troodons were
much larger than their more southern counterparts.
 |
| Artist: Maija Karala Ugrunaaluk |
Among the polar fossil finds are remains
of fern, cycad, conifers and pollen from shrubs and herbs. This was truly a
self-sustaining biodiversity. All the fossils were found in the 70
million-year-old ‘Lipscomb’ fossil bed that is a part of the 80-60 mya Prince
Creek Formation, made up of alluvial deposition layered on top by mud flats.
These mud flats give evidence of multiple dinosaur tramples alluding to herding.
Down at the other end of Earth in what
is now the Antarctic, from Cretaceous rock oxygen isotope levels, the estimated
mean annual temperatures were between 0 and 8 °C/32 and 46 °F. Fairly warm for
today’s Antarctic subzero temperatures but still cold for what we think
dinosaurs could thrive in. Rocks the dinosaur fossils were found, in an area
called ‘Dinosaur Cove’ give evidence of permafrost from features such as ice
wedging and hummock ground. Permafrost normally occurs in temperature ranges of
between −2 and 3 °C/28 and 37 °F. Also, there were indeed long wintry polar
nights with periods of up to six months of total darkness, although there is no
evidence of a polar ice cap. The Antarctic floral community at this period
consisted of forests of conifers, ginkgoes, ferns, cycads, bryophytes,
horsetails and even a few flowering plants.
In the Early Cretaceous, East Gondwana
was composed of Antarctica still linked to Australia after both had rifted and
drifted from Africa. Much of East Gondwana’s southern portion at the time laid
inside the Arctic Circle. It is in this region where most of the Antarctic and
Australian dinosaur fossils are found. By the end of the Cretaceous, Australia
had fully rifted apart from Antarctica moving further away from the South Pole
and Antarctic Circle, taking away any land bridge. This separated the Antarctic
and Australian dinosaur species from their respective kin, which then evolved
into differing genera.
Antarctica had six genera of dinosaur
with four herbivores in the ankylosaur Antarctopelta
(An-tarc-toe-pel-tah) and the three ornithopods, Atlascopcosaurus
(At-lass-cop-ko-sawr-us, Morrosaurus (Moe-row-sawr-us) and Trinisaura (Tree-nee-sore-rah);
one omnivore in, Glacialisaurus
(Glay-see-al-ee-sore-us) and one carnivore in, Cryolophosaurus (Cry-oh-low-foe-sawr-us). For the Australian
portion that was once a part of Antarctica, nineteen genera fossils have been
discovered.
 |
| Artist: Solec Kujawski Atlascopcosaurus |
Of the Antarctica dinosaurs, the basal ornithopod,
Atlascopcosaurus showed up 114 mya in
the Early Cretaceous. It was ~ 2-3m/6.5-10ft long and most likely, as other
iguanodontids was horn beaked and could move either on the two longer hind
limbs or on all fours. On an interesting note, this is the only dinosaur to be
named after the Swedish corporation, ‘Atlas Copco’ who manufactures
construction and mining equipment and also assisted in the excavation of the
fossil remains.
 |
| Artist: Mike Belknap Antarctopelta |
As a herbivorous quadruped, the
nodosaurid, Antarctopelta occurred in
Antarctica during the Late Cretaceous 74-70 mya. At 4m/13ft, it had osteoderms
(armored bony plates embedded into the skin with shoulder and neck spikes and a
short spike extending from the supraorbital bone that pointed outwards from below
the eyes.
 |
| Artist: Sebas Runa Morrosaurus |
 |
| Artist: Peter Trusler Trinisaura as the cold caught up |
 |
| Artist: Peter Trusler Glacialisaurus eating from tree |
 |
| Cryolophosaurus skeletal anatomy |
 |
| Artist: Joe Tucciarone Cryolophosaurus |
 |
| Artist: Fabio Pastori Lft:Cryolophosaurus Rt:Dilophosaurus |
Besides chasing down herbivorous
dinosaurs, we also know Cryolophosaurus
dined on other animals, as the mammal-like reptile cynodont, tritylodont’s
teeth were found in the gut cavity of Cryolophosaurus’
fossil remains. But also, the remains showed tooth marks randomly put of other
smaller theropods that most likely were consuming Cryolophosaurus’ carcass.
Dinosaur
Clades:
There is a world of dinosaurs and to go
over each one would require an exposé in encyclopedic form. Just in sauropods
alone there are over 120 known species. So what we’re going to do is discuss
sauropods, ornithischians and theropods in the respective group levels finishing
with the theropod lead-in into birds.
All dinosaur lineages evolved from within a single group of archosaurs such as the proterosuchids and Euparkia, which was elaborated on under the pterosaurs of ‘Et Tunc Nulla Erat VII’. Even though dinosaur radiation included carnivorous bipeds and herbivorous quadrupeds, they all arose from bipedal omnivores with long grasping hands such as Nyasasaurus (243mya), and herrerasaurs 230-220 (mya).
 |
| Simplified cladogram on the path to dinosaurs |
All dinosaur lineages evolved from within a single group of archosaurs such as the proterosuchids and Euparkia, which was elaborated on under the pterosaurs of ‘Et Tunc Nulla Erat VII’. Even though dinosaur radiation included carnivorous bipeds and herbivorous quadrupeds, they all arose from bipedal omnivores with long grasping hands such as Nyasasaurus (243mya), and herrerasaurs 230-220 (mya).
Sauropodomorpha:
Just a reminder...all sauropods are
sauropodomorphs, but not all sauropodomorphs are sauropods. This is the case
for any infra ordering of related species as the refinement of clans become
more specific. To bring this to terms in thinking about it, all Homo species are primates, but not all
primates are Homo species.
 |
| Sauropodomorpha Clade |
Sauropodomorphs are the long necked
herbivorous dinosaurs that existed from the Late Triassic to the Late
Cretaceous 231.4-66 mya. The long necks fit an eating niche in accommodating
greater access to higher up tree limbs. They became the dominant herbivore
before the end of the Triassic and managed to survive the extinction events
between the Triassic/Jurassic boundary and the Jurassic/Cretaceous boundary.
For the most part, except for the earliest species, they were quadrupeds. The
latter forms developed a four legged graviportal (slow moving over terrain)
gait due to their heavier size and length.
 |
| Artist: Nobu Tamura Saturnalia |
Although smaller at first, sauropodomorphs
later became the largest terrain animals to ever walk the earth. Of 228 mya, Saturnalia (Phonetics: Sah-tur-nay-lee-uh)
was only 1.5m/5ft long while the massive Argentinosaurus
(Ar-jen-teen-oh-sawr-us) was 35m/115ft that could tremble the ground on every
step 97-93.5 mya. Although only an estimate, due to lack of fuller fossil
material, the argentinosaur species, Argentinosaurus
huinculensis may have reached a length of 39.7m/130ft. Sauroposeidon (Sawr-o-pss-sy-don) from 120 mya was probably the
tallest, reaching about 18m/60ft high. Reaching extremities in size was a
natural selection sequence for defense, for other than size they only had weak
teeth and a retained thumb with a claw on their forelimbs for defense.


The teeth, shaped like spoons weren’t even efficient for masticating food, so the swallowing of stones (gastroliths) for grinding and relying on bacteria to break down the nutrients evolved within the sauropodomorph’s digestive strategies. They also possessed large nares (nostrils) and had 25 presacral vertebrae just like in most humans that have 23-25 presacral vertebrae.
 |
| Artist: Cisiopurple Sauropodomorphs |
The most derived sauropodomorph
discovered thus far is, Panphagia (Pan-fah-eg-ah)
and Chromogisaurus (Kro-mo-ge-sawr-us)
that lived 230+ mya. Panphagia was a
mere 1.3m/4.3ft while bigger, but still really small for sauropods, Chromogisaurus was 2m/6.7ft. Both were
bipedal and in the process of transitioning from carnivores to sauropod
herbivores, these sauropodomorphs were omnivores and insectivorous. All
primitive and basal sauropodomorphs went extinct after the Triassic/Jurassic
extinction.
 |
| Lft ~ Artist: Nobu Tamura Panphagia Rt ~ Artist: Cisiopurple Chromogisaurus |
 |
| Herrerasauridae Cladogram |
Some of the earliest dinosaurs come from
the family Herrerasauridae
(Phonetics: Huh-reah-ruh-soar-uh-day). Herrerasaurid dinosaurs like the Late
Triassic, 235-228 mya, Caseosaurus (Kay-see-o-sawr-us)
and the 235-210 mya, Chindesaurus (Chin-dee-sawr-us)
carried anatomy traits possessed by all three dinosaur subgroups making them
very difficult to classify in whether belonging to theropods, ornithischians or
sauropods. All herrerasaurids were bipedal carnivores. The ~231.4 mya, Herrerasaurus (Huh-reah-ruh-sawr-us)
possessed derived archosaur traits, but also basal dinosaur traits. As a basal
trait, the ilium was supported by just two sacrals, while as a derived trait,
its pubis pointed backwards.
 |
| Credit: UK Nat'rl His. Museum Chindesaurus |
 |
| Artist: Mark Hallett Staurikosaurus |
 |
| Artist: Gonzalezaurus Sanjuansaurus |
 |
| Artist: Apsaravis 2 Herrerasaurus & a rauisuchian |
Three herrerasaurids in, Herrerasaurus, Sanjuansaurus (San-wan-sawr-us) and Staurikosaurus (Store-ick-o-sawr-us) are relegated as a node clad
polytomy group, meaning more than two descendants are involved within a
cladogram and in this herrerasaurid case, placed outside of saurischian’s
direct taxonomic ancestry. But, these three herrerasaurids are still listed as
the most primitive group of Sauropoda.
 |
| Artist: Julius T. Csotonyi Aardonyx |
A 190 mya Early Jurassic sauropodomorph
showing the evolutionary transition into sauropods was, Aardonyx (Ard-o-nix). Representing the transition from biped
sauropodomorphs to quadruped sauropods, it was bipedal itself, but could easily
walk as a quadruped. With the pelvic girdle and hind limb structuring, it was a
biped, but with stiffening rigidity taking over flexibility in the forelimbs,
it was trending towards a quadruped weight bearing form.
 |
| Artist: Heidi Richter Massospondylus |
One of the oldest dinosaur fossil embryo
is from the 4m/13ft in length sauropodomorph, Massospondylus from the Early Jurassic 200-183 mya. The 190 mya
fossil embryos were from a nest site in what is now South Africa. The developed
and well preserved embryos were still in the eggshells and at birth would’ve
only been 20.3cm/8in long.
 |
| Artist: Julius T. Csotonyi Massospondylus |
Sauropoda:
As a subgroup to sauropodomorphs, sauropods
are the giants we immediately think of when contemplating dinosaurs. All
sauropods were herbivores and developing their digestive strategies as
explained previously, they required heavier bodies to accommodate the bulk load
of food and the size of its digestive systems in the gut areas.
 |
| Sauropod Cladogram |
In concentrating on bigger trending
torsos, sauropods basically retained ancestral head sizes making it
proportionally smaller as compared to the evolving large body. Most sauropod
fossil discoveries are minus the head and it’s due to the small skull being
easily carried or washed away from the rest of the skeletal remains. But an
exceptional sauropod skull discovery of titanosaurian, Sarmientosaurus (Sar-me-in-toe-sawr-us) gives us some glimpses on
what this sauropod depended on.
 |
| Artist: Marcos Paulo Sarmientosaurus skull/head |
The skull measured only 43.2cm/17in, but
was attached to a 12.2m/40ft long dinosaur. With all the skull cavities intact,
CT scans of the skull showed the brain was the size of a plum, but the scans
also detailed sarmientosaurs had highly developed visual and auditory sensory
reception; more so than other sauropods. Large eye sockets and long ear canals
show that this sauropod had acute vision and was able to hear distant low
frequencies. Also, the shape of the skull and neck arrangement shows that the
head was held downwards in life suggesting Sarmientosaurus
would sweep through low foliage and vegetation.
To support the heavier body, columnar elephantine
same sized quadrupedal limbs evolved, but not like an elephant’s limb distal in
splaying outwards creating a wide foot. Sauropod feet had the manus bones
arranged in fully vertical columns, with extremely reduced finger bones.
Sauropods walked in digitigrade pes fashion where their heel and metatarsals were
lifted off of the ground. In eusauropods though, there is exception.
Eusauropods were the bigger sauropods with skeletal adaptations to support the
weight and lengths utilizing asymmetry in their metatarsal shaft diameters
where the thumb metatarsal is broader than the others, shifting the load
bearing weight onto the inner half of the feet.
 |
| Artists: Raul Martin/Kristina Curry Rogers Rapetosaurus |
Sauropod young looked like a miniature
adult. This gives credence that the young were on their own in finding food and
were not being fed by the parents.
 |
| Artist: Dinoraul Mamenchisaurus |
It was the sauropod body that affected
growth rates resulting in an increased rate of growth. In order to achieve this,
some type of metabolic warming had to occur. Sauropods most likely weren’t
endothermic, but with their enormous digestive system giving off heat, it’s
highly probable they were linked to tachymetabolic endothermy. Tachymetabolism
maintains a high ‘resting’ metabolic rate and even though sauropod ‘resting’
metabolism is many times slower than their ‘active’ metabolism the switch was
permanently on in generating heat. This is a form of heterothermy as seen in
today’s bats and small birds where the animal can switch from poikilothermy to
homeothermy and back again.
Sauropods are known for their long
necks. The longest necked sauropods were the mamenchisaurids and a recent 2015
China fossil discovery of Qijianglong
(Chee-jee-yahn-long) had one of the longest mamenchisaurid necks. Filled with
air sacs, sauropod necks were light weight as compared to neck size. The long
necks also contained a long trachea for the lungs to gain more oxygen. From
fossil analyses, sauropods neck anatomies experienced, ‘Osteological Neutral
Pose’ (ONP) where all the neck vertebrae are neutrally articulated with no deflection
up or down. In other words the neck was not disarticulated enough to give
efficient sideways motion.
As in today’s giraffes, sexual
dimorphism is with the male in having larger necks. Females are more attracted
to males with the largest necks. It can be proposed that this scenario as well
was with sauropods, as male sauropods indeed did have the bigger necks and it
is with this neck dimorphism that over millions of years of time probably led
to the most derived sauropods in being the ones with the largest necks.
 |
| Artist: Greg Paul sauropod necks |
With long necks and heavy bodies, the
tails were also long for counterbalance. The tail also may have been used for
defending off predators and using them as a whip snapping the air quick enough
to break the sound barrier. This could have been utilized as a warning signal
to group members of approaching predators, or to distract carnivores.
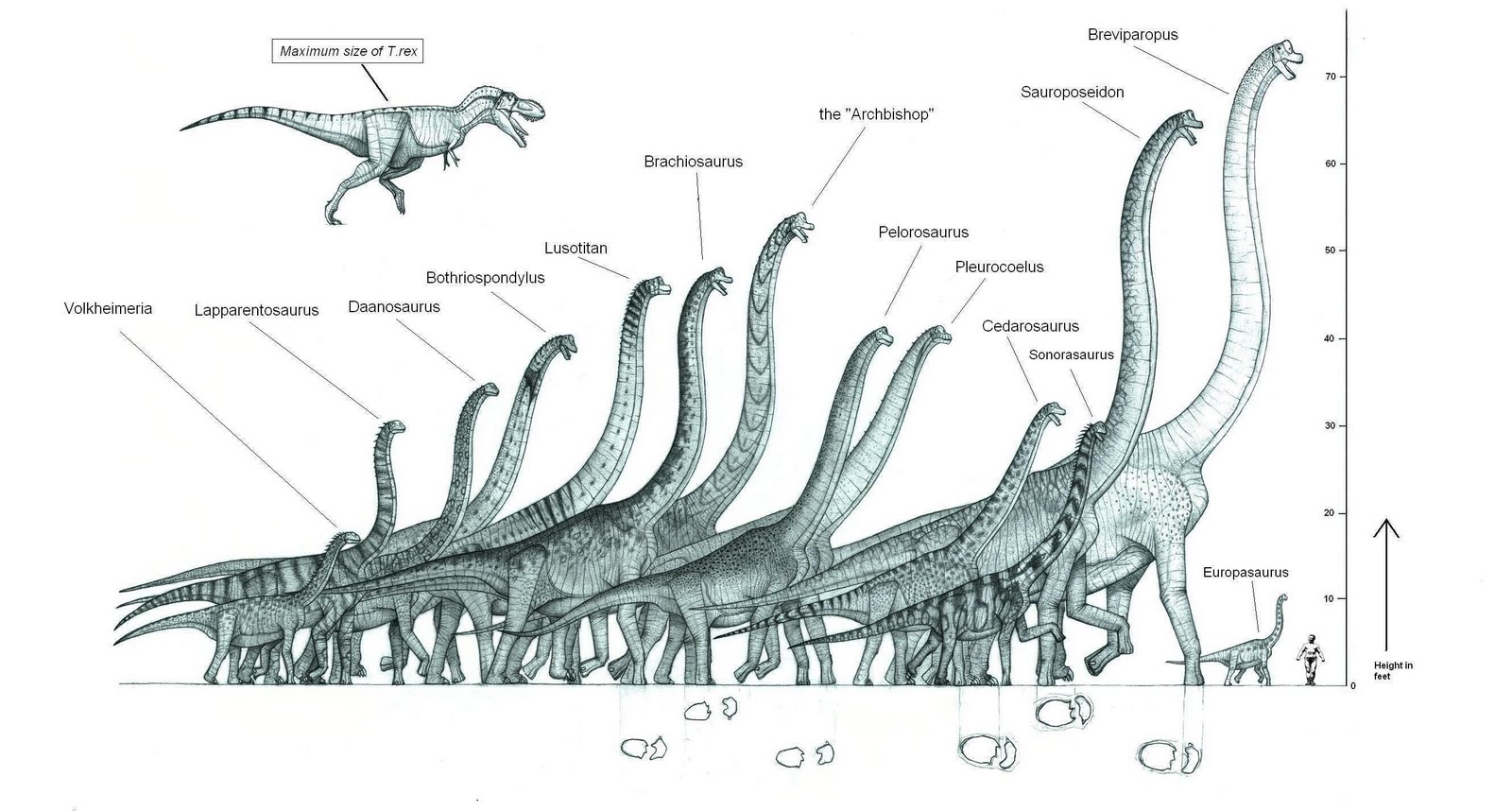 |
| Artist: Paleo King With long necks then long tails for counterbalance |
It appears that most sauropods stayed in
small bands and clustered with other sauropod species. In the Jurassic, there
were nine major subgroups with numerable minor groups composed of numerous
genera within each that coexisted. How in the world did they not over compete
for food resources and make certain groups lose out to extinction? First off,
sauropods, in addition to having spooned or spatula like teeth for grasping and
taking in food, other forms evolved another type of dentition of peg-like
teeth. This alludes to the fact that varying sauropod families preferred
different types of plant foods.
 |
| Sauropod dentition |
Microscopic studies on the wearing of
teeth shows coarsely scratched and pitted wear on the spoon types and finely scratched
surfaces on the pegged teeth. In comparing microscopic studies of extant
herbivorous mammals and in knowing what they ate, it gives credence to what
sauropods long ago ate. Camarasaurids, with the spoon teeth ate coarser plant
material, whereas the diplodocids, with the more evolved pegged teeth had the
more finely scratched dentition.
This advocates that sauropods ate selectively;
one preferring softer foods such as aquatic plants and shoots, while the other
consumed tougher shrubs and older plant parts that had collected wind-blown
dust grains. It also shows eating behavior patterns as peg-like teeth would
work in nipping off shoots or water plants, where spoon-like teeth were far more
adequate in grabbing whole mouthfuls of plant material, like leaves and stems. But
with apatosaur, nigersaur and rebbachisaur, among others, studies displaying
large proportional pitting, fine subparallel scratching dentition and along
with blunted square snouts, these sauropod groups likely were nonselective low to
the ground grazers. So, sauropods covered browsing behaviors from ground height
to mid-level and to upper canopies.
 |
| Artist: Steveoc 86 Melanorosaurus |
Melanorosaurus (Meh-lan-or-o-sawr-us)
lived during the Late Triassic 227-221 mya. Some articles will list Melanorosaurus as a true sauropod, but it
is not. It was a most derived sauropodomorph that was however, an intermediate
between advanced sauropodomorphs basal to primitive sauropods. It most
certainly had sauropod features such as the transitioning to a large sauropod
neck bauplan. This of course accelerated an access to a food source that was
out of reach for most other herbivores of the day. The extensive pneumatization
of the sauropod axial skeleton was also evident in Melanorosaurus as inherited by sauropods. This paved the way for
sauropod evolutionary advantages in lowering the energy cost for pulmonary
breathing, reducing specific gravity of its mass and would aid in eliminating
excess body heat.
Although the forelimbs were slightly
shorter than the hind limbs the femur was straight, thick and columnar sitting
immediately below the body, although the pes and manus distal metacarpals were
disarticulated splaying from one another as in their more primitive
predecessors. With true sauropods, the toes and fingers were encased in flesh
and not visible; only the claws were. This made for a stronger padded round base
in supporting weight.
A common feature Melanorosaurus had with other more primitive sauropodomorphs was in
having nine premaxillary teeth, but had up to nineteen maxillary teeth that were
common in a lot of the more derived sauropods. This animal was ~10.7m/35ft and
although this is small as far as more derived sauropods became to be gigantic,
it most likely was the largest animal during its day.
Antetonitrus (An-tee-toh-nigh-trus) lived during the Late Triassic 210 mya. It is a basal sauropod and although it easily walked as a quadruped, the forelimbs were shorter and not as adapted for bearing weight. Antetonnitrus etymology is Latin in ‘ante’ (before) and ‘tonitrus’ (thunder) and is in reference to Brontosaurus (Bron-toe-sawr-us) which is Greek meaning, ‘thunder lizard’.
Antetonitrus (An-tee-toh-nigh-trus) lived during the Late Triassic 210 mya. It is a basal sauropod and although it easily walked as a quadruped, the forelimbs were shorter and not as adapted for bearing weight. Antetonnitrus etymology is Latin in ‘ante’ (before) and ‘tonitrus’ (thunder) and is in reference to Brontosaurus (Bron-toe-sawr-us) which is Greek meaning, ‘thunder lizard’.
 |
| Artist: Karkemish 00 Antetonitrus |
Digit I (first digit or thumb) of the
forelimbs was set higher up from the other digits and was twisted 40° making it
opposable to the rest of the foot. The manus as a whole wasn’t locked into
position inferring digit I was used for grasping. In judging the forefeet, Antetonnitrus was the most basal
sauropod in the transitional phase from bipedal sauropodomorphs to true
quadrupedal sauropods. Quadrupedalism requires a pronated manus so that it may
swing in a parasagittal plane. The distinctive tri-radiate proximal ulna of Antetonitrus and derived sauropods may
be linked to a medial shift in the position of the radius that brought about
this pronation. Other than the forefeet anatomy, this sauropod had already
evolved typical sauropod features of the digestive tract, long necks/tails and
columnar bearing limbs directly below the body.
 |
| Artist: Fabio Pastori Bilkanasaurus |
Blikanasaurus (Bly-khan-ah-sawr-us)
was another one that teetered between being designated as a sauropodomorph
doomed to a dead end, or to be classified as one of the oldest sauropods thus
found. But after recent studies, the conceding final analysis is that it is the
most basal sauropod thus discovered, although not as a direct ancestor to
behemoth sauropods that were later to come.
Occurring 225 mya in the Late Triassic,
as Antetonnitrus, Blikanasaurus had the opposable digit I,
but in analysis of its skeletal structure, was one of the first sauropods to
have a permanent quadrupedal gait even though digit I was not weight bearing.
Occurring 110 mya toward the end of the
Early Cretaceous was, Brontomerus (Brawn-toe-meh-rus). The name is Greek meaning, ‘thunder thighs’ and
for very good reasoning. This sauropod had the largest and strongest thigh
muscles of any animal thus far known. The huge muscle projected forward. Even
the ilium on which the thigh muscle was attached projected forwards suggesting
the ability to deliver a strong forward kick. The ilium further had attachments
for abductor muscles allowing the leg to draw up and away from the body. For
this roughly, 60 metric tonnes/66 US tons (132,277 lbs.) sauropod with a length
of 14m/46ft, the strong hind limbs assuredly gave it advantages not afforded to
other sauropods.
 |
| Artist: Francisco Gasco Brontomerus kicking Utahraptor |
The capable strength and power delivered
in this kick definitely could have served a fatal blow to any predator of its
day. It may have also served in males kicking it out during mating periods or
standing side-by-side kicking one another for leadership dominance. The hind
limb musculature also would give Brontomerus
an advantage of greater support in going up or down steep inclines and a
steadier base for rearing up on the hind limbs. In being able to climb and
descend steep hills and to rear safely on its hind limbs gave a greater access
to plants that would be unattainable for other large sauropods.
 |
| Artist: Tumas Koivurinne Shunosaurus |
Living in the Middle Jurassic 170 mya, Shunosaurus (Shu-no-sawr-us), with 21
China fossil finds, was one of the most unusual sauropods. At 9.5m/31ft, it was
fairly small for evolving sauropods and had the second shortest neck among
sauropods, just behind the shortest neck of Brachytrachelopan
(Brak-e-trak-el-o-pan). Brachytrachelopan’s
neck is 40% shorter than other sauropods, where Shunosaurus is 37% shorter. This suggests they were low browsers.
 |
| Shunosaurus tail fossil |
Another Shunosaurus oddity is its upper and lower jaws curved upwards that
functioned much like a pair of garden shears in stripping foliage. But the
oddest trait was its mimic of ankylosaurs in having the tail end in a bony club
topped by two 5cm/2in osteoderm cone-shaped spikes used for fending off
predators and perhaps male competitors when mating.
Rapetosaurus (Phonetics:
Rah-pay-to-sawr-us) is thus far one of the last sauropods that held out to the
end of the Cretaceous 77-66 mya. It is under the clade, Titanosauria, but as under that order, was very moderate in size at
15m/49ft in length; being half the size of other titanosaurs. Juvenile remains
have also been found with its length at 8m/26ft. As pointed out and pictured earlier
in a few paragraphs above, paleontologists also discovered an infant Rapetosaurus with its size no more than
a medium sized dog. Implementing bone histology and X-ray computerized
tomography, the researchers found that the infant had the same ratio of bone
compactness per size as the adult alluding to the fact that Rapetosaurus resembled the adult stage
throughout all the sub adult stages, indicating a very early independence from
parental care. The small size could be referred to island dwarfism as Rapetosaurus was isolated on the forming
island of Madagascar that rifted from Africa in the Early Cretaceous 135 mya and
India during the Late Cretaceous 88 mya.
 |
| Artist: Tyler Keillor Rapetosaurus weeks old |
Rapetosaurus also shared
similar skull features of Diplodocus
(Phonetics: Dih-plaw-duh-cuss) in possessing a more slender cranium with the
nares (nasals) opening up between the eyes. It also had closely quartered
pencil-like dentition more suited for stripping leaves away from tree branches.
As a born and raised West Texan, I have
to speak of Glen Rose, Texas. The Paluxy River runs through the quaint town and
outside a few miles away, dinosaur tracks were discovered in the riverbed. Way
back as a child in the late 1950s and early 1960s my siblings and I would
scamper down into the riverbed during the dry season checking out the dinosaur
trackways and reliving what it would be like to actually see one of those
dinosaurs that left those massive limestone foot impressions. It let our
imaginations run wild. Today though, that part of the river is Texas’ ‘Dinosaur
Valley State Park’.
 |
| Old Paluxy Poster |
 |
| Theropod track in Paluxy River bed |
 |
| Dino tracks Paluxy River |
 |
| Artist: Cheung-tat Chung Acrocanthosaurus pursuing Paluxysaurus |
Anyway, in honoring those childhood
days, I give you the Paluxy River’s, Paluxysaurus
(Pa-lux-ee-sawr-us). Up to 18m/59ft long there is a portion of a Paluxysaurus
foot path that was excavated along the Paluxy River’s bedrock now housed in New
York’s, ‘American Natural history Museum’. The Paluxysaurus tracks are trailed by a large theropod predator, which
was most likely left by Acrocanthosaurus
(Ak-row-kahn-tho-sawr-us), the most prevalent carnivore of that Early
Cretaceous time period. Paluxysaurus
lived 115-110 mya and Acrocanthosaurus,
116-110 mya. Although according to its skeletal remains, Acrocanthosaurus wasn’t very speedy. The foot trails depict Paluxysaurus being tailed by a theropod
of Acrocanthosaurus’ 11.5m/38ft size
range. Paluxysaurus remains were
found in Hood County in which Glen Rose is the county seat, while Acrocanthosaurus fossil remains have
been found throughout Texas near Hood County. The fossil footprints and
impressions may not have been left by Acrocanthosaurus,
but the evidence retained in them certainly holds a vast amount of info of that
animal.
 |
| Artist: Linda Bucklin B. excelsus |
On a closing note, as but a small child
I always equated long necked dinosaurs as a brontosaur. Then as an adult, it
was deciphered that the brontosaur fossils were actually just another apatosaur.
I so grieved, for I missed my brontosaur. But alas, as an older man there is to
be no more grieving as it has been positively borne out through meticulous
research that not only is there a brontosaur, but there are actually three
species in, B. excelsus (the very one
that was removed and reassigned as an apatosaur species then back to brontosaur),
B.
yahnahpin, and B. parvus.
Below are a few sauropods:
 |
| Artist: James Kuether Apatosaurus |
 |
| Artist: Luis V. Rey Amargasaurus |
 |
| Artist: Luis V. Rey Astrodon fending off pesky Utahraptors |
 |
| Artist: Phil Wilson Brachiosaurus |
 |
| Artist: James Kuether Omeisaurus |
 |
| Artist: Kostyantyn Ivanyshen Titanosaurus |
Ornithoscelida:
Besides the common bird-hip,
ornithoscelidans were also intermediate in bird structure with various
ornithischian species possessing integumentary filaments, quill-like appendages
and feather-like epidermal branching structures. Of course in theropods,
distinct evidence in ulnar quill knobs, down feathering and feathers have been
found in their fossil remains. In both of these dinosaur groups there is also
fossil evidence of epidermal scales, just as it is present in bird legs today. So
for dinosaurs comprising, Ornithoscelida, scales and feathering
were evident in skin covering as a whole, or in part or in possessing both.

 |
| Ornithoscelida simple clades |
This new dinosaur cladistic relationship
grouping also suggests that hyper-carnivory was acquired independently in herrerasaurids
and theropods, offering an explanation for many of the anatomical features
previously regarded as notable convergences between theropods and early
ornithischians. The earliest theropods, ornithischians and also sauropodomorphs
to arise, basically came from the same grandparents; it’s just that theropods
and ornithischians were first cousins and second cousins to sauropods in matter
of terms.
Ornithischia:
First off, please don’t confuse ‘Ornithoscelida’ with ‘Ornithischia’, for it’s easy to do. They
are as distinct from one another as Australopithecus
is to Homo sapiens. One covers multiple variant groups (Ornithoscelida) while the other (Ornithischia) carries a few specific related subgroups.
 |
| Ornithischia Cladogram |
Ornithischians came in many various
forms as quadrupeds and bipeds, as armored or bony, but what primarily
designates a dinosaur to be related to the ornithischian club is the pelvic
anatomy. Ornithischian pelvises, or hip girdles have the pubis and ischium
bones running parallel to one another pointing backwards just below the ilium
which points forwards. This is extinct and extant bird hips’ typical
arrangement, thus the name, Ornithischia
which is Greek for ‘bird hipped’. They also possessed ossification on the
predentary of the dentary (lower jaw) that formed a beak extending the dentary in
coinciding with the upper premaxillary.
Other ornithischian characteristics were
toothless paired premaxillary bones with a keratinous tipped beak and possessed
a palpebral bone that laid across the outside of the eye socket (similar to an
eyebrow function in humans). Also, these dinosaurs shared a much reduced or
closed off antorbital fenestra (skull openings in front of the eyes) and
stiffened backbones near the pelvis due to ossified tendons above the sacrum.
All ornithischians were herbivores.
Those with teeth bore leaf-shaped, or
premaxillary spoon-shaped dentition or a battery of peg-like teeth set in back
of the mouth that was surrounded by rhamphoteca (keratinous sheaths).
Ornithischians with dentition chewed softer plant material inaugurating the
digestion process. The keratinous beaked ornithischians with few or no teeth
relied on a large gut for bacterial fermentation.
Some, like the ceratopsid, Psittacosaurus (Phonetics: Sit-tah-coe-sawr-us),
the ankylosaurid, Panoplosaurus (Pan-op-low-sawr-us),
the iguanodontid, Iguanodon (E-gwan-o-don)
and the hadrosaurid, Claosaurus (Clay-o-sawr-us)
had gastroliths found in the gut area of their fossil remains, but other than
these four dinosaurs, gastroliths are not evident in ornithischian fossils. To
induce proper digestion in assimilating nutrients from plant bulk, most
ornithischians may have incorporated gastric milling. This is a process where a
musculature stomach is equipped with a layer of hard keratin and small pebbles
which breakup, pulverize and emulsify food content. Toothless ostriches today
incorporate the same process. As ornithischians are related to theropods and
birds evolved from theropods, it is conceivable herbivorous birds genetically
inherited gastric milling from ornithischians. Found at the front of the lower
jaws, all ornithischians possess a predentary bone that functions as an aid in
cropping plants.
Ornithischians are divided into two
major subgroups; they are: Thyreophora
(Thyr-uh-ree-off-for-uh) and Cerapoda
(Sair-uh-po-duh). The clade Cerapoda further
divides more derived ornithischians into two subgroups, Ornithopoda (Or-nith-op-o-duh) and Marginocephalia (Mar-gin-o-seff-fol-e-uh). Cerapods were the bird
foot dinosaurs where marginocephalians were the ‘bone headed’ and ‘horned face’
dinosaurs. The dinosaurs that make up these three clade groups are:
Thyreophora: ankylosaurs,
stegosaurs.

Ornithopoda: hadrosaurs,
hypsilophodontids, iguanodontids, thescelosaurids, jeholosaurids, yandusaur.

Marginocephalia: A) Pachycephalosauria ~ pachycephalosaurs;
B) Ceratopsia ~ Yinlong, Chaoyangsaurus, psittacosaurids,
ceratopsids.

Although the Late Triassic suchian, Revueltosaurus (Re-vul-toe-sawr-us)
living 225-209.5 mya was originally thought to be a basal ornithischian, it
appears that ornithischians did not arise until the Jurassic, as revueltosaurs
were finally determined to be a clade of suchian archosaurs.
 |
| Revueltosaurus skeletal anatomy |
Fabrosaurs, agilisaurs, nanosaurs and
gongbusaurs occurring throughout the Jurassic 199-155 mya ago, have scant
fossil finds and cannot be reliably classified, but at what has been found and studied,
they are primitive bipedal ornithischians. Ranging from 1-2m/3.3-6.6ft in size,
unless perfectly fossilized, their fragile remains were easily disintegrated
throughout the eons of Earth upheavals and weathering.
 |
| Artist: Wayne Barlowe Fabrosaurus |
After much debate, Pisanosaurus (Pi-san-o-sawr-us) has been determined to be the oldest
ornithischian thus far discovered despite the fact it is not a true dinosaur,
but a dinosauriform. Although it is not a true cerapod, it does show ornithopod
anatomical features with affinities to hypsilophodontids and heterodontosaurs
and is the earliest known bird-hipped animal. Living 228-216.5 mya in the Late
Triassic, Pisanosaurus was a biped
herbivore reaching a length of 1m/3.3ft. The acetabulum (hip-joint) is open,
low and axially long, due to the short pedicels of the ilium. The upper region
of the ischium is wide and larger than the pubic bone. Pisanosaurus is the sister group of the basal ancestor to heterodontosaurids
and thyreophorans.
 |
| Artist: Masato Hattori Pisanosaurus |
Occurring in the Early Jurassic 199-189
mya, Lesothosaurus (Leh-so-toe-sawr-us)
was only 1m/3.3ft long, but with good fossilization more is known of this
ornithischian. Lesothosaurs are one of the first true dinosaurs to have evolved
the bird-hip. The hind limbs were long in proportion to body size while the
forelimbs were short, but with four well developed metacarpals (finger bones).
The anterior end of both the lower and upper jaws was a characteristic
ornithopod-like keratinous beak for snipping off plant material. Inside the
mouth in the front were canine-like teeth followed by leaf-shaped teeth in the
lower jaws that meshed with the horny beak in an up and down motion to shear
food. There is evidence that lesothosaurs traveled in small herds.
 |
| Artist: Albert Gruvitz Lesothosaurus |
Heterodontosauridae (Hay-tear-o-don-toe-soar-uh-day)
species are the most basal members of ornithischians. Occurring from the end of
the Triassic through the Jurassic and on into the Early Cretaceous 201-133 mya,
the heterodontosaurid family consisted of three genera and one subfamily, Heterodontosaurinae composed of five
genera as illustrated in the cladogram below.
 |
| Heterodontosaurid Cladogram |
 |
| Credit: NRF S. Africa Abrictosaurus |
 |
| Heterodontosaurid dental evolution |
 |
| A) Heterodontosaurus B) Tianyulong |
 |
| Heterodontosaurid fossil with feathering integuments |
One of the striking features found in
heterodontosaurid fossils are the impressions formed from filamentous
integumentary structures on the back, tail and neck, which are a theropod
variant of protofeathers. Also, heterodontosaurid forelimbs were lengthening; a
trending factor towards quadrupedalism.
One unique anatomical characteristic found in at least some, like the pigeon-sized Manidens is in the feet. The pes (foot) had a raised ridge on the back near the ankle and is interpreted as an area for additional tendon attachment. The toes were unusually long ending in narrow and curved claws resembling many birds. Finally, the first toe had a unique combination of characters suggesting that it may have been able to grasp objects despite being shorter than the other three toes. This comes to the conclusion that like most perching birds; some heterodontosaurids could clamber and perch upon tree limbs.
One Late Jurassic 150 mya heterodontosaurid was Fruitadens (Phonetics: Fru-tye-dens). Measuring only 65-75cm/25-30in in adult length, it was a small ornithischian. This diminutive dinosaur was also the only ornithischian omnivore as evidenced by its dentition and no, it did not eat fruit. The name comes from the locale its fossils were found in, which is near the town of Fruita, Colorado. Remember, fruiting plants didn’t come about until a good 20 million years later in the Cretaceous. With hind limbs as long as its head and torso, it was a swift runner.
 |
| Artist: Julio Lacerda Heterodontosaurid perching |
One unique anatomical characteristic found in at least some, like the pigeon-sized Manidens is in the feet. The pes (foot) had a raised ridge on the back near the ankle and is interpreted as an area for additional tendon attachment. The toes were unusually long ending in narrow and curved claws resembling many birds. Finally, the first toe had a unique combination of characters suggesting that it may have been able to grasp objects despite being shorter than the other three toes. This comes to the conclusion that like most perching birds; some heterodontosaurids could clamber and perch upon tree limbs.
 |
| Artist: Masato Hattori |
One Late Jurassic 150 mya heterodontosaurid was Fruitadens (Phonetics: Fru-tye-dens). Measuring only 65-75cm/25-30in in adult length, it was a small ornithischian. This diminutive dinosaur was also the only ornithischian omnivore as evidenced by its dentition and no, it did not eat fruit. The name comes from the locale its fossils were found in, which is near the town of Fruita, Colorado. Remember, fruiting plants didn’t come about until a good 20 million years later in the Cretaceous. With hind limbs as long as its head and torso, it was a swift runner.
 |
| Artist: Tyler Keillor Pegomastax |
One other heterodontosaurid was even
smaller at 60cm/2ft. Occurring 200-190mya in the Early Jurassic, Pegomastax (Peg-o-mass-taks) was an
herbivore that utilized its dentary fangs to uproot plants, dig for roots and
as a secondary use in defense. In possessing the paired fangs, porcupine-like dorsal
quills and extremely long legs for sprinting, this dinosaur was defensively
equipped no matter its size.
A few more heterodontosaurids:
A few more heterodontosaurids:
 |
| Artist: Nobu Tamura Tianyulong |
 |
| Artist: Luis V. Rey Heterodontosaurus tucki |
 |
| Artist: Nobu Tamura Manidens |
Thyreophora: Thyreophorans
comprise the ankylosaurs and stegosaurs; the armored dinosaurs. Their temporal
range was from the Late Jurassic to the Late Cretaceous 199.6-66 mya. Utilizing
etymology, Thyreophora is derived
from the Greek ‘thyreos’ (shield) and ‘phoros’ (bearer), in reference to their
various types of armor. Instead of feathering, thyreophorans opted for heavy
ossification and osteoderm skin covering. Except for the basal thyreophoran, Scutellosaurus (Skuh-tell-o-sawr-us),
all other thyreophorans were quadrupeds walking on all fours with shorter
forelimbs than the hind limbs.

Basal or primitive thyreophorans had not
yet developed the armored osteoderm plates that the more derived forms, like
stegosaurs and ankylosaurs are known for, but from fossil studies of preserved
soft tissue, it is concluded that most basal thyreophorans possessed an outside
layer of keratinous skin. From this, the armor derived in the later forms.
The most primitive thyreophoran evidence
thus found are, Tatisaurus (Tah-tee-sawr-us)
found in China’s Lufeng Formation of Early Jurassic sedimentary rock dated
199.5 mya and Bienosaurus (By-no-sawr-us).
Bienosaurus was also found in the
same ‘Lufeng Formation’ as Tatisaurus
and is estimated to be from 195+ mya. Both of these finds are skimpy and Tatisaurus is considered to be nomen
dubien due to the fossil only being a dentary (lower jaw) with teeth still
intact. But the teeth and jaw have enough traits to tie it into its ornithiscian
fabrosaur ancestry and thyreophoran lineage. Bienosaurus’s fragmentary skull and jaw remains have close
affinities to the thyreophoran, Scelidosaurus
(Scale-e-do-sawr-us).
 |
| Artist: Luis V. Rey Scelidosaurus |
Occurring 196 mya in the Early Jurassic,
Scutellosaurus is one of two of the
most basal thyreophorans that led to the more derived armored forms. Scelidosaurus, from 191 to 183 mya of
the Early Triassic, is the other. Both were strict herbivores. With only a
maximum length of 1.2m/3.9ft, Scutellosaurus
was bipedal, but with longer hind limbs, the shorter forelimbs were still long
and built strong enough to perform weight bearing qualities, so it could walk
in a quadrupedal stance with ease. It possessed osteoderms that were fixed as
plates of bone within the skin. The osteoderms ran dorsally down the back and
flanks in parallel rows. This would have given Scutellosaurus an advantage from early theropods of the day in
protecting it from teeth piercings of the skin and internal organs. Of course
more derived theropods evolved into larger forms and stronger bites, so
thyreophorans in kind evolved into larger sizes with more armor. Scutellosaurus had a long tail that was
likely used as a counter balance to the heavier armored body.
 |
| Artist: James Kuether Scutellosaurus |
Closely related to Scutellosaurus, Scelidosaurus
was a notch above in the armory. Still retaining the shorter forelimbs, Scelidosaurus was fully quadrupedal and
possessed six rows of osteoderm bony plates down the dorsal and flank sides of
the body, with the back four dorsal rows ending at the tail tip. In addition,
there were three pointed plates just behind the skull. A well preserved Scelidosaurus fossil find showed preserved soft tissue around the vertebrae that was an
epidermal layer of keratinous skin overlaying the osteoderms. At 4m/13ft long, Scelidosaurus was nearly triple the size
of Scutellosaurus and is the sister
taxon to the two main thyreophoran clads of stegosaurs and ankylosaurs
 |
| Credit: BBC Co. UK Scelidosaurus fossil |
Representing an immediate form between Scutellosaurus and Scelidosaurus, the 4m/13ft long Emausaurus
(Em-ow-sore-us) from the Early Jurassic 184-176 mya, had a skull anatomy that
was very similar to Huayangosaurus, (Hue-way-ang-o-sawr-us).
The Emausaurus skull was also small
compared to its 4.5m/14.8ft body that was also to become a distinguishing
feature of stegosaurs.
 |
| Artist: Graham Rosewarne Emausaurus |
The two major thyreophoran clads are the
stegosaurs and ankylosaurs. The clad, Stegosauria
covers fourteen species and Ankylosauria
covers six species. Stegosaurians are known for their rows of bony upright
spikes or plates running down their spines while the more armored
ankylosaurians are more known for patches of external bones fused into
protective plates. Stegosaurs’ temporal range was from the Middle Jurassic to
the Early Cretaceous 165-136 mya. Ankylosaurians had a longer temporal range
from the Middle Jurassic to the Late Cretaceous 167-66 mya. Ankylosaurs were
one of the last groups of dinosaurs to go extinct.
 |
| Artist: Franco Tempesta Huayangosaurus |
 |
| Hesperosaurus varying gender plates |
 |
| Artist: Masato Hattori Hesperosaurus mjosi male |
Huayangosaurus was a basal
stegosaurian found in China and as such, Asia is the origin of stegosaurians.
Coming from the Middle Jurassic 165-163.5 mya, Huayangosaurus is a basal stegosaurian and was small at 4.5m/15ft
in length when compared to the more derived forms. It also possessed from its
ancestral line premaxillary teeth that were disposed of in later forms. Called
‘thagomizers’, it was the first stegosaur to possess these spikes (thagomizers)
on the end of its tail which were two paired extending outwards horizontally
and sideways. In addition to having the rowed paired of bony plates running
down its back, above its hips in the plate rows were paired spikes. As for the
plates themselves, they were narrow and tipped as opposed to later stegosaurian
plates being more rounded. With a low stance and broad skull, Huayangosaurus was a low browser and did
not have adaptive capabilities for a specialized diet.
 |
| A) Correct B) Wrong Gigantspinosaurus |
 |
| Artist: James Kuether Gigantspinosaurus |
Occurring around 160 mya, its plate rows
were also narrow and tip pointed just as Huayangosaurus,
but ended just before the tail. Huayangosaurus
and Gigantspinosaurus had a unique
feature that was absent in all other subsequent stegosaurians in having
ossified tendons extending down the vertebral column.
Stegosauridae is the family
of stegosaurs within the suborder of Stegosauria.
It’s defined by all the stegosaurians having a closer relationship to
Stegosaurus (Steg-o-sawr-us). Stegosaurids roamed in the Middle Jurassic to
Early Cretaceous 165-136 mya. The vast majority of stegosaurians so far recovered
belong to Stegosauridae with most
species arising from the Late Jurassic and Early Creataceous, but by the latter
part of the Early Cretaceous there was a decline in diversity and speciation.
There was a fossil find in India coming from the Coniacian Stage (89.8-86.3
mya) of the Late Cretaceous that was once thought to be a stegosaur dubiously
named, Dravidosaurus, but turned out
to be a plesiosaur. The decline and final extinction might be due to a floral
turnover when angiosperms began to dominate, drastically reducing cycadophytes which
corresponded with the stegosaurid decline. As it stands, Wuerhosaurus (Where-ho-sawr-us) at 5m/16ft long living ~ 136 mya in
the Early Cretaceous, was the last of the stegosaurs to go extinct.
Stegosaurids were evolving wider hip placements and Wuerhosaurus, as one of the last stegosaurids had the widest hips.
This is thought to be the result of enlargement of the digestive tract.
Stegosaurids also possessed flexible armor-like scales that protected the
throat.
 |
| Artist: Masato Hattori Wuerhosaurus |
 |
| Artist: Vladimir Nikolov Kentrosaurus |
 |
| Artist: Mohamad Haghani Miragaia |
Miragaia (Meer-uh-guy-uh)
as a stegosaurid, also retained the shoulder spikes that were long and pointed
at the tips; although there is current debate if the spikes were from the
shoulders or actually were a specialized tail spike. In addition, this odd
stegosaurid with a temporal range of 150 mya in the Late Jurassic had unique
rows of eight paired plates that were tipped like a picket wooden fence transitioning
down into the body’s dorsal side asymmetrically with a convex outer side and a
concave inner side. The plates’ base weren’t as expanded as other stegosaurids
except for the last narrower pair. The plates ran down the spinal column from
the back of the head to the rump. Beyond the rump and down the tail, the plates
were transformed into spikes.
The most unique attribute of Miragaia however was its neck. Through a
Hox gene switch trigger, it’s theorized these regulatory genes that organize
organism body plans, once activated from dormancy, induced back vertebrae to be
carried forward becoming neck vertebrae. As a result, Miragaia had 17 neck vertebrae giving it the longest neck of all
other stegosaurians, even more than most sauropods, by almost doubling the neck
vertebrae count in the other stegosaurid members, even though more derived
stegosaurs were evolving lengthened necks. It may never be truly clear what
natural selective pressures were lengthening stegosaurid necks and in
particular Miragaia’s neck. It could
be an advantage to reach higher for food sources in utilizing the tail and hind
limbs in a tripod stance since its body’s center of gravity was more towards
the hips. Or, it was to use the neck in a swaying back and forth motion to
vacuum up lower plant material in swoops. Or it could even have been a sexual
attractant to the opposite gender. With the lengthened neck, Miragaia’s total length was 6m/29ft.
One other thing on Miragaia, its remains were found in Portugal where most stegosaurid
fossils have been found in North America. This only verifies continental drift
as Miragaia’s temporal range was when
N. America was connected to Western Europe.
 |
| Artist: Karen Carr Stegosaurus |
Stegosaurus (Steg-o-sawr-us)
in modern culture is the famous dinosaur that the family, Stegosauridae is named after. Living in the Late Jurassic 155-150
mya, this stegosaur was a low browser feeding on soft vegetation. Its bite
force was less than a human’s due to weak jaw muscles, so it nipped off soft
vegetation with its turtle-like beak by pulling the head back. It then relied
on its 78 small triangularly flat teeth to chew the food thoroughly before
swallowing, allowing the enlarged gut aided with gastroliths to do the main digestion
of nutrients. It did indeed eat well though for its total adult length was 9m/29.5ft
in length, making it the largest of all stegosaurids.
Stegosaurus had a
graviportal gait in that it was adapted to moving slowly. This was not due to
dimwittedness. An excellent fossil find of a well preserved stegosaurus
braincase showed that the brain cavity held a brain weighing no more than
80g/2.8oz. This is about the size of two walnuts. Very small, for a 4.5 metric
ton/5 short ton animal, but the graviportal gait was solely due to a high body
weight as a result of the long hind limbs and not due to brain size. There were
three species of stegosaurs in S. stenops, S. ungulatus and S. sulcatus.
Stegosaurs had two pairs of spikes at
the end of its tail that pointed outwards for defense. The main stegosaur predators
were allosaurs. Paleontologists have found allosaur fossils with puncture
wounds that stegosaur thagomizers (tail spikes) would perfectly fit into.
A few more stegosaurs below:
 |
| Artist: Alain Bénéteau Lexovisaurus defending against Allosaurus |
 |
| Artist: Cisiopurple Stegosaurs |
 |
| Artist: James Kuether (Paleoguy) Tuojiangosaurus defending against Yangchuanosaurus |
Ankylosauria ~ The sister
group to stegosaurians, were ankylosaurians, the tank of dinosaurs. I
personally prefer referencing ankylosaurians as the dinosaur equivalent to
armadillos. Standing low to the ground, they were not fast runners with short
stocky limbs and as armadillos, their ventral side or underbelly was void of
any armored protection as the rest of the body was; just like armadillos.
Armadillos protect the vulnerable underbelly by curling over it, while
ankylosaurians mastered the low to ground stance under a heavy and broad
armored body to protect it; differing methods fashioned toward the same end result.
 |
| Ankylosauria clade |
Ankylosaurians were low browser feeders
and due to whatever foliage was available in ecosystems of their biomes,
evolved differing methods for assimilating food. As borne out through tooth
wear and mandible articulation studies, some groups chewed with tooth occlusion
(teeth contact) accompanied with palinal (backward motion) jaw movement. Other
groups chose a widening and shortening of the muzzle evolving a convergent precise
tooth occlusion, while others evolved the same process aided by an additional
palinal power stroke with a narrowing and elongation of the muzzle. In the more
basal groups of Asia, processing food was restricted to orthal (vertical motion
effects) pulping. These types of nutrient assimilation were evolutionary
responses to what flora was presented, as with only an inflexible neck reach of
no more than one meter ankylosaurians had to depend on whatever foliage was
available near the ground. However, the hind limbs were capable enough in
holding the body weight if the dinosaur wanted to support itself on a trunk
with the forelimbs to get at low hanging branches.
The enameled phylliform (leaf shaped)
teeth as triangular shaped, were similar to stegosaurians and were replaced
when worn out. Possessing hyoid bones, suggests they had long flexible tongues
to wrap around soft food plants and had a large secondary palate indicating
they were able to still breathe while chewing.
Distinguished features of the heavily
ossified ankylosaurian skull shows that this group evolved early by splitting
off into its two main families of, Nodosauridae
(No-doe-sawr-uh-day) and Ankylosauridae
(An-kill-o-sawr-uh-day). The ankylosaurian tail evolved one step at a time from
the stiffened nodosaurid tail to the ankylosaurid tail ending in bony clubs.

Nodosauridae: Nodosaurids were
the first of the ankylosaurians to appear 155 mya in the Late Jurassic,
becoming extinct 66 mya at the end of the Cretaceous. Nodosaurids are considered
to be more related to Panopolosaurus
(Pan-op-lo-sawr-us) than to the ankylosaurids. All nodosaurids had neck and
flanked body spikes projecting outwards sheathed over with keratin, except for Panopolosaurus where spikes were absent.
All nodosaurids had heavy osteoderms and bony nodule rows running dorsally down
the back and sides of the head and body. Unlike as their ankylosaurid kin, the
tail ended in a typical dinosaur tapered, but flexible tip with no armaments. The
snouts were relatively narrow and triangular shaped indicating a preference to
selective browsing of low growth plants.

Nodosaurids had a global distribution,
including Antarctica in the genus, Antarctopelta
first mentioned above under Dinosauria.
They all became extinct by the end of the Cretaceous, but it was not an abrupt
event due to the asteroid impact; it was more of a gradual decline in family
members.
 |
| Mymoorapelta skeletal anatomy |
Living 155-150 mya in the Late Jurassic,
Mymoorapelta (My-moor-ah-pel-tah) is
the most basal nodosaurid currently discovered and was found in Colorado, USA. The
oldest known nodosaurid from Europe is from the Early Cretaceous 112 mya named Europelta (Your-o-pel-tah). It had some
odd traits in having the ratio tibia to femur lengths greater than any other
ankylosaurian, possessed autapomorphic (a derived trait unique to one species)
osteoderms located on the outer corners of the pelvic shield and the skull was
covered by a single osteoderm. With fossil remains found in Austria, the last
nodosaurid to go extinct was, Struthiosaurus
(Stru-the-o-sawr-us) living in the Late Cretaceous 85-66 mya. At 2.2m/7.2ft
long, it was also the smallest of the ankylosaurians.
 |
| Artist: Roman Garcia Europelta |
Nodosaurids fossil remains have been
discovered in near shore marine sediment topologies suggesting that they
preferred littoral coastal plain habitat, with some carcasses washed out to
sea. Ankylosaurid fossil remains have only been found inland in more upland
habitats.
 |
| Artist: David Bonadonna Borealopelta drowning |
As described more like a mummified
dinosaur or a statue of one, the fossil remains coming from an open pit oil shale
mine in Alberta, Canada is one of the most well preserved fossils ever to have
been discovered. In fact, the whole body is in place giving a 3-D image of a
sleeping animal. The nodosaurid, Borealopelta
(Bor-e-al-o-pel-tuh) occurred ~ 110 mya near the end of the Early Cretaceous.
The fossil was found in marine sediment and was washed out to sea in the
Cretaceous landlocked shallow ‘Western Interior Seaway’.
 |
| Where Borealopelta was found N.G. |
Studying the pigments in the fossil’s
preserved skin, it’s been determined the body coloration was a reddish brown in
a counter-shaded pattern most likely for camouflage. Unfortunately, before the
miners spotted the fossil the posterior portion was pulverized into unusable
debris by heavy digging equipment.
 |
| Borealopelta's fossil |
Below a few more nodosaurids:
 |
| Artist: W. Parsons Tatankacephalus |
 |
| Nodosaurids |
 |
| Artist: Masato Hattori Hylaeosaurus |
Ankylosauridae: Where ankylosaurids
don’t have long spikes on the neck and shoulders, but have a tail ending in a
weaponized club, nodosaurids had long spikes on the neck and shoulders but a
clubbed tail was absent. Found only in the Northern Hemisphere of N. America,
Europe and Asia, ankylosaurids lived in the Early Cretaceous 122 mya until the
Late Cretaceous 66 mya.

The ankylosaurid clubbed tail first evolved by weaving caudal vertebrae into bulbous bundles. As the mapped out evolutionary history of ankylosaurid tails portrayed, earlier ankylosaurids bear out the gradual fusing of the tail vertebrae making it less flexible in successive speciation. This essentially turned the tail into a stiffened rod enacting it to work as a handle. If the tail had still been flexible, it would not have supported the weight of the club, potentially tearing tendons and muscle. Also, the stiffened tail, before the club evolved, most likely was already being used for defense by swinging it into the legs or torso of an oncoming predator; much like a nightstick or billy club. The knobbed club was formed by osteoderms fusing in layering bone within the skin at the tip of the tail.
 |
| Ankylosaurid skull |
Ankylosaurids had shorter limbs than
nodosaurids which would make it harder for a predator to overturn in getting at
the non-armored underbelly. With massive skulls, ankylosaurid heads were as
broad as long and were shaped as triangular with dermal scutes/plates solidly
ossified to cartilage bone. The spine was rigid and curved permanently arching
the back.
 |
| Credit: Australia Geographic.co Kunbarrasaurus |
Considered as basal to ankylosaurids was
the ankylosaurian, Kunbarrasaurus (Coon-bah-rah-sawr-us)
formerly known as ‘Minmi’, but now
has been designated its own genus. It lived 119-113 mya during the Early
Cretaceous. The fossil remains were found near the region in Australia’s ‘Minmi
Crossing’ in the Allaru and Bungil Formation made up of sandstone, siltstone
and mudstone sediment. The cranial endocast (brain casing) is similar to later
more derived ankylosaurids, but other head features were very unique. It had
more of a tuatara-like proportionally enormous inner ear and a nasal airway
that looped back in on itself, where ankylosaurids had straw-like nasal
passages. Gut cavity fossil remains consisted mainly of fibrous plant tissue
fragments from twigs and stems, with lesser amounts of seeds and fruiting
bodies swallowed whole. The chopped fibrous material and lack of gastroliths
suggest that food was snipped with the beak and masticated thoroughly before
swallowing.
 |
| Artist: Eloy Manzanero Chuangqilong |
Chuanqilong (Shwahn-chee-long)
lived during the Early Cretaceous ~ 110 mya in what is now China and is
considered one of the earliest basal ankylosaurids. The most derived ankylosaurid
and last to go extinct was Ankylosaurus
from 66 mya. It was also the largest ankylosaurid reaching lengths of
8m/26ft.
 |
| Credit: Ji Q., Wu X., Cheng Y., Ten F., Wang X, Ji Y. Lianpningosaurus |
All ankylosaurians were strict
herbivores. Well that is absolutely true except for one major exception; the
ankylosaurid, Liaoningosaurus (Lao-nin-go-sawr-us)
indeed ate plants, but supplemented its diet with at least fish making it also
a piscivore. This Early Cretaceous ankylosaurid, living 122 mya had partially
digested several fish remains found within fossilized gut cavity. It was found
in China’s Liaoning Province in the ‘Yixian Formation’ that is made up of
volcanic basaltic rock and siliciclastic volcanic material. The fossil was of a
complete sub adult that either scavenged or hunted fish. There is fossil
evidence that it was at least semiaquatic with longer leg ratios to body as
compared to other ankylosaurids and there was lack of hip fusion allowing more
flexibility in hind limb paddling. Also, the tail was flexible enough for
sinusoidal propelling, found in no other ankylosaurian stiffened tail fossils.
It also had a flat single osteoderm covering the ventral (belly) portion of the
body. This would have protected the underside from predators attacking from
below as it swam in freshwaters. However, there is now discussion on whether
the belly plate is a true bony osteoderm or preserved skin.
Below are a few more ankylosuars:
 |
| Artist: Sydney Mohr Above: Gobisaurus Below: Ziapelta |
 |
| Artist: Cisiopurple Ankylosaurids |
 |
| Artist: Jake Baardse Ankylosaurus |
Cerapoda: As mentioned
above under Ornithischia, cerapods
are divided into two groups: the ornithopods and the marginocephalians. Under
marginocephalians, cerapods are further divided into two subgroups in the ornithopodans
and ceratopsians. Cerapods arose during the Middle Jurassic 169 mya and went
extinct in the Late Cretaceous 66.5 mya.
 |
| Simple Cerapoda Cldogram |
 |
| A bit more detailed Cerapoda cladogram |
Cerapods transitioned and radiated out
throughout the Jurassic and into the Cretaceous. The main cerapod success was
in their dental adaptations evolving ways to thoroughly extract maximum
nutritional value through intensive mastication of plants that were otherwise
too tough and rough for other dinosaur digestive tracts. Cerapod species were
the first to assimilate thorough digesting limited primarily to chewing. It
wasn’t until much later that mammalian herbivores would take up this role.
The term, Cerapoda combines its two subgroups with ‘cera’ (horned) from ceratopsians
and ‘poda’ (foot) from ornithopodans.

Ornithopoda: There were
three distinct family clades of ornithopods belonging to the families, Hypsilophodontidae (Phonetics: Hip-sill-loff-o-dawnt-uh-day),
Iguanodontidae (E-gawn-o-dawnt-e-day)
and Hadrosauroidea (Had-row-saw-roi-dee-uh)
were ornithopods with a temporal range from the Early to Late Cretaceous 130-66
mya. Ornithopods were bipedal but occasionally could maneuver a quadruped gait.
These groups were low browsers grazing on plant roughage that was easily
cropped and masticated due to the evolving dentary assemblage and chewing
mechanisms. It was the most sophisticated form of assimilating food than any
other herbivorous non-avian dinosaurs. Later forms of ornithopods developed
curved spines allowing them to graze on all fours.
 |
| Artist: Oscar Sanisidro Hypsilophodon |
Hypsilophodontidae: This family
group is now being considered paraphyletic as the last descendent form from its
common ancestor. Where before other genera were included, it now only includes
the one genus species, Hypsilophodon
foxii (Hip-sill-loff-o-don = fox-eye). Although this small ornithopod
(1.8m-5.9ft) occurred well into the Early Cretaceous 130-125 mya), Hypsilophodon had very primitive
ornithischian features much like Pisanosaurus
and may have been omnivorous. Like the common opossum, Didelphis marsupialis, there were never enough environmental
pressures to further stress evolvement.
 |
| Artist: Mark Hall Oryctodormeus |
Close ornithischian relatives to Hypsilophodon, were two burrowing
dinosaurs in, Drinker (Phonetics:
Drin-ker) occurring ~ 150 mya in the Late Jurassic and Oryctodromeus (Phonetics: Oh-rik-toe-dro-mee-us) occurring nearly
100 million years later in the Late Cretaceous 95 mya. Named after pioneering
paleontologist, Edward Drinker Cope,
Drinker was a primitive basal relationship to Hypsilophodon and while Oryctodromeus
was related to Hypsilophodon, it was
also basal within the line of ornithopods.
These two herbivorous biped swift
runners were around 2m/6.6ft long and had modified forelimbs, broad snouts,
flexible tails, slender pelvises and shoulder girdles. These adaptations represent
a burrowing lifestyle. In fact, there was a family of Oryctodromeus in a fossil find that had a parent and young. The
collapsed burrow portion of the fossil discovery had solidified previously dug
up loose dirt and was filled with sand that turned into sandstone as a stark
contrast from the formation’s ambient mudstone and claystone. The den was much
like hyenas excavate today.
Added burrow advantages were safety
from predation and extreme temperatures. In addition, in finding the family
huddled together, the fossil find points to parental care. With that in mind, both parents in this scenario
were taking care of the young. We can only imagine the scene this fossil
represents of this cave-in with the frantic behavior of the one parent outside
realizing there was nothing it could do to extricate the family from being
buried alive.
 |
| 100 mya honeybees amber fossil |
 |
| Iguanodontid manus (hand) fossil |
 |
| Monophyletic Iguanodontid Clade |
Iguanodontids were monophyletic in that
they descended from a common ancestral line not shared with any other
ornithopod group. Tenontosaurus (Ten-non-toe-sawr-us),
occurring 115-108 mya was the most basal iguanodontian. In fossil finds, the
presence of medullary bone was found in a thigh and shin bone that for today is
only found in female birds when producing eggs.
 |
| Artist: H. Kyoht Lutterman Muttaburrasaurus |
An ornithopod trending towards the
iguanodontid form was, Muttaburrasaurus
(Phonetics: Mut-tah-buh-rah-sawr-us) that occurred ~ 105.8 mya in the Early
Cretaceous was basal to iguanodonts and is a member of an iguanodontid clade.
Its skeletal anatomy is close to that of iguanodontids, but it was incapable of
quadrupedalism and had teeth for slicing and shearing instead of the
iguanodontid battery of teeth for chewing.
In taxonomic nomenclature, the term, Iguanodontoidea (Ig-wan-o-don-toy-dee-uh)
is a ‘superfamily’ ranking that is not as specific as ‘family’ phylogenies. Whenever
in dealing with animal taxonomy the suffix, ‘oidea’ refers to a superfamily. As
an iguanodontoid, in today’s terms means it could be an iguanodon, but not
quite yet there. This is the case for, Barilium
(Ba-ril-e-um) that occurred 140 mya in the Early Cretaceous. Given its
divergent phylogenetic position, it’s been separated out from the Iguanodon genus. It was a very stocky,
even chubby dinosaur set with large vertebrae featuring short stubby neural
spines to support the body weight. The hind limbs were almost elephantine with
the distal pes covering a broad surface area.
 |
| Artist: Mark Witton Barilium |
Above is paleontologist, Mark Witton’s
interpretation of Barilium with
folded and thick leathery skin; a contrast in dinosaur interpretations in
always having smooth skin. The animals below Barilium are not birds but tiny maniraptoriform dinosaurs.
 |
| Artist: Mark Witton Iguanodon |
A derived iguanodontid form was Iguanodon (Ig-wan-o-don) with a temporal
range of 126-125 mya during the Early Cretaceous. Growing as long as 13m/43ft
in length, Iguanodon was also bulky
at 3.08 tonnes/3.4 tons in weight. The forelimbs were 75% the length of the hind
limbs and walked both as a biped and quadruped as evidenced by fossilized
footprints. Tendons were ossified along the neural arches facilitating both
modes of pedal movement in absorbing most of the stresses incurred in
bipedalism. This ossification limited backbone mobility in exchange for
reinforcement. All of the cervical vertebrae had the ribs attached.
 |
| Iguanodon fossil |
In the forelimb manus (hands), they were
almost as useful as primate hands. Digits II, III and IV were close together
where III and IV may have been bound together. Digit I (thumb), as opposable,
was essentially a spike. Nevertheless, the manus would have been capable of
grabbing. There can only be conjecture on what the spike was used for. It could
have been used for gouging in defense from predation and intraspecific
competition, or for males positioning the females during mating, or for all
three conjectures. Skull muscle attachments suggest it had a large tongue.
 |
| Artists: Ray Moller/Dorling Kindersley manus (hands) |
Iguanodontids rapidly evolved into
hadrosauriforms leading to hadrosaurids. The transition was so rapid on a geologic
scale that Lurdusaurus’s
(Lur-du-sawr-us) snout was becoming duck-billed, while Altirhinus’ (Al-ti-rain-us) snout was hollowing.
A few iguanodontids below:
 |
| Artist: Sergey Krasovskiy Altirhinus |
 |
| Artist: Elperdido Lurdusaurus |
 |
| Credit: wiki.com Tenontosaurus |
Hadrosauridae: Called the
‘duckbilled dinosaur’ due to the snout shaped similarly as a duck’s bill, did
not possess any type feathering as other ornithischians had as all the
innumerable hadrosaur skin impression fossil finds show only scalation of the
skin. The snout rostral bones were flattened, thick and laterally compressed.
There are three subfamilies of hadrosaurids being: Hadrosaurinae (Had-row-sore-e-nye), Saurolophinae (Sore-o-lop-phee-nye) and Lambeosaurinae (Lam-bay-o-sore-e-nye). Assumed for mating or alarm calls,
lambeosaurines possessed hollow cranial crests/tubes made from the premaxilla
and nasal bones that would sound once the animal exhaled through the nostrils.
Hadrosaurids are considered facultative bipeds with most young walking
bipedally while adults were mainly quadrupeds. Jaw design was to grind
vegetation to a pulp aided by multiple rows of teeth that were replaced once
worn down. Although solid crests were found in some of the species two
subfamilies, most hadrosaurines and saurolophines did not support cranial
crests; only lambeosaurines had hollow crests used for bellowing.

 |
| Artist: Julius T. Cstonyi Ouranosaurus |
 |
| Ouranosaurus skelatal anatomy |
 |
| Artist: Willydynamo Maiasaura nesting |
 |
| Artist: Brian Switek Maiasaura hatchlings |
 |
| Artist: Oliver Volland Maiasaura parental care |
Maiasaura (May-uh-sawr-uh) literally means ‘good mother’ in reference to a fossil find composed of Maiasaura eggs, embryos and young found in a nesting colony. This supports parental care in at least the mother feeding the young in gathering fruit and soft plants. Mothers were too large to sit on nests without breaking the eggs, so as crocodilians, placed fermenting vegetation on top of the nest to provide heat as the plant material rotted. In rookeries, chances of egg and young survival were improved with the amount of gathered adults.
 |
| Artist: NestieBot Shantungosaurus |
The largest hadrosaurid was Shantungosaurus (Shan-tun-gui-sawr-us)
with a skull size of 1.63m/5.3ft and length of 16.6m/54ft. The beak was
toothless but the jaws were packed with ~ 1,500 teeth made for chewing. A large
hole near the nostrils is found in the fossils and is conjectured to have been
covered by a loose flap of skin for bellowing. Shantungosaurus temporal range was in the Late Creatceous 78-74
mya.
 |
| Artist: Mihai D. Dumbrava Telmatosaurus |
A few more of hadrosaurids:
 |
| Artist:Masato Hattori Mukawaryu recently discovered 2013 |
 |
| Artist: Luis V. Rey Charonosaurus |
 |
| Artist:Victor Leshyk Gryposaurus |
 |
| Artists: L. Xing/Y. Liu Lambeosaurus |
 |
| Artist: Luis V. Rey Various hadrosaur heads |
Marginocephalia: Dinosaur groups under Marginocephalia were all herbivores existing from 156-66 mya. Some were the last of the dinosaurs to live just prior to the Cretaceous/Paleogene extinction, but all became extinct during and just after the event. These ornithischians as grouped into the two subgroups, pachycephalosaurians and ceratopsians are best known as the ‘bone head’ pachycephalosaurians and the ‘horn faced’ ceratopsians, like the three horned Triceratops (Tri-sair-ah-tops). But, Triceratops was just one species of many dinosaurs under the family, Ceratopsidae (Sair-ah-top-see-day). There are three subfamilies under Ceratopsidae and at least forty-four genera. Pachycephalosaurians were bipeds and are the one herbivorous dinosaur group that never trended towards quadrupedalism. The earliest basal ceratopsians and Psittacosaurid ceratopsians were facultative bipeds, while all other ceratopsians were quadrupeds, with the exception of the bipedal, Archaeoceratops (Ar-kay-oh-seh-rah-tops). The predentary and rostral supported a horny beak in all marginocephalosaurians.

Marginocephalians first evolved in the
Late Jurassic 156 mya and thrived until the end of the Late Cretaceous 65.5 mya,
thus being one of the last groups of dinosaurs to go extinct. All the earlier
forms, or primitive marginocephalians originated in what is now the Asian
continent, with the later and more derived migrating up into what is now North
America.
Marginocephalians were social creatures
and through fossil evidence roamed in small to large herds and formed rookeries
for their eggs and hatchlings. They also herded with other dinosaur groups.
There is evidence as well of interspecies communication implying cooperative
communication between other species and with this sharing and understanding,
formed a mutualism factor that benefitted all species concerned. This spurs the
idea that interspecific communication in the transfer of information, whether
by language or sight signaling is yes, first a learned trait, but is through an
evolutionary mechanism.
Pachycephalosauria ~ Living at the
end of the Early Cretaceous to the end of the Late Cretaceous 99-66 mya,
pachycephalosaurians are known for their thickened cranial skull caps. There is
debate on proper mobility, but they were bipeds and could casually walk on all
fours at times such as grazing on low vegetation. However, pachycephalosaurians
are the one herbivorous dinosaur group that never trended towards
quadrupedalism. Most pachycephalosaurians were small, averaging 2-3m/6.6-9.8ft,
but as most small dinosaurs were, they were not climbers, instead were limited
to accessing food no higher than one meter above the ground.

In terms, the order, Pachycephalosauria and the family, Pachycephalosauridae were studied and
reevaluated in 2006 and from that, it was determined that they both included
the same species taxa. As a result, it was concluded that Pachycephalosauria was redundant and struck its ordering leaving
only Pachycephalosauridae to list all
pachycephalosaurians and pachycephalosaurs into the one family group. But this
groups all less bony flathead and bonier dome head pachycephalosaurs into one
family. I disagree with that; but who am I, right? Anyway, since the flatheads
were more primitive and older than the more derived bone heads, along with
other anatomical, even physiological differences, I feel that the group, Pachycephalosauria and its immediate
basal ancestors should remain and that is why I’m presenting it as such. Just
to inform ya, so enough stated...
 |
| Artist: Cisiopurple Wannanosaurus |
I’m listing Wannanosaurus (Wan-nan-oh-sawr-us) here as a primitive
pachycephalosaurian basal to pachycephalosaurids. In today’s literature it most
likely will be listed as a pachycephalosaurid. Occurring in the Late Cretaceous
80 mya, Wannanosaurus was a tiny at
60cm/24in and supported a flattened skull with large fenestrae. The fossil find
gives evidence of fused bones determining that it was an adult. It may have
also supported some dorsal quilled feathering. According to dentition, it most
likely was not a strict herbivore as later pachycephalosaurids were, but was
omnivorous in dieting on plant material and insects.
Pachycephalosauridae: Due to
combining both pachycephalosaur groups into one, this family contains eighteen
genera of pachycephalosaurs, but due to the uncertainties in the polarities of
species characters, makes for a less cladistic analysis. There is also ongoing
debate as to if the flatheads were simply subadult forms of the domed boneheads.
The temporal range was in the Late Cretaceous 76-65.5 mya. The revised
definition of pachycephalosaurids is among other traits, those
marginocephalosaurians with thickened but fully flat to fully domed
frontoparietals (frontal and parietal bones of the cranium), the squamosal was a
deep plate on the occiput having enlarged upper-outer corners, the premaxillary-maxillary
diastema was arched and possessed developed rows or clustered nodes or blunt
horns on the squamosal, nasals and in species specificity, other parts of the
skull.
Concerning the domed boneheads, it was
first interpreted that the thickened skull was for intraspecific combat in head
butting or for interspecific defense against predators. This idea was bolstered
through dome head cranial histological studies where 22% of all domes examined
had healed lesions consistent with osteomyelitis, an infection of the bone
resulting from penetrating trauma. In flatheads there was 0.00% of any head
trauma, which supports the subadult flathead thesis as mating combat is
performed only by adult males.
The bone tissue of the domes was
composed of a unique form of fibrolamellar material containing fibroblasts that
play a critical role in bone healing. This further accelerates the proponents’
idea of pachycephalosaurid ‘agonistic behavior’ defined as any social behavior
related to fighting. Also, to aid in absorbing the head-butting blows, the
spinal column possessed vertebral articulations providing spinal rigidity.
However, pachycephalosaurids carried their necks in U or S shaped curves and
could not step back and charge each other with straightened necks spread out
horizontally for head on butting.
 |
| Artist: De Agostini |
Pachycephalosaurids’ head, neck and
spine could not align itself in such a way as to absorb, alleviate, or transmit
stress like an extant musk ox or mountain goat can today. In addition, domed
head butting would for the most part result in glancing blows due to the
curvature. This would’ve created much more damage to the head due to the nodes
and clusters of small horns (spikes) creating open wounds to the domes’ skin or
keratinous coverings and other parts of the head that would have created
serious infections. As well, to reduce weight, the fibrolamellar material was
spongy and would crush under the force of head on butting.
So, I support the side that feels,
although pachycephalosaurids weren’t head bangers, they did indeed use the
domes for intraspecific ‘flank butting’ that would allow the domes to give a
bruising side blow to an opponent’s hindquarters, but still not do too much
damage to the head. For sure the domes evolved as weaponry and not as species
identification, for all pachycephalosaurid domes were indistinct in anatomy and
appearance.
 |
| Artist: Nobu Tamura Homalocephale |
 |
| Credit: dinoweb.narod.ru Goyocephalae |
 |
| Dracorex skeletal anatomy |
 |
| Artist: Darren Horley Dracorex |
 |
| Artist: Vlad Korstantinov Pachycephalosaurus |
The last two pachycephalosaurs to go
extinct were the pachycephalosaurids, Pachycephalosaurus
and Sphaerotholus (Sfay-ro-fo-luss)
with a Late Cretaceous temporal range of 70-66 mya and 73-66 mya respectively. Both
are considered the most highly derived of all pachycephalosaurians. While the
three genera of Sphaerotholus lengths
are unknown due to lack of body fossil material, size probably averaged no more
than 2.4m/7.9ft. As for Pachycephalosaurus,
it so far is the largest pachycephalosaur at 4.5m/14.8ft.
 |
| Artist: FishPainter Sphaerotholus |
Some pachycephalosaur pics:
 |
| Artist: Julius T. Cstonyi Acrotholus |
 |
| Artist: Dinoraul P. wyomingensis |
 |
| Credit: Paleo-World Prenocephale |
 |
| Artist: Cisiopurple Various pachysaurs |
Ceratopsia ~ Although ancestral forms lived as far back as 161.2 mya in the Late Jurassic, the bulk of ceratopsians lived during the Cretaceous Period. As in pachycephalosaurians, the main feature in ceratopsians was in the skull’s bony ornamentation. Ceratopsians eventually embellished on the skull ornamentation, but instead of bony domes and small spikes, they elaborated on frills and horns.

Unique to ceratopsians, the rostral bone
located on the tip of the upper jaw anteriorly joins the two premaxillae
forming a single midline bone. The modification represents an upper jaw mirror
image of the predentary and like the predentary the rostral also supported a
horny beak. The jugal skull bone pointed laterally to form a horn-like process.
Essentially there were two main
evolvement influences within ceratopsians during the Cretaceous. The first
evolution sequence involved psittacosaurs (121-99 mya) giving rise to Protoceratops (Pro-toe-sair-ah-tops) 83
mya. In turn, Protoceratops gave rise
to Styracosaurus (Sty-rah-co-sawr-us)
then to Triceratops (Tri-sair-ah-tops)
67 mya.
Ceratopsians are further divided into Neoceratopsia, Leptoceratopsidae, Ceratopsoidea
and Ceratopsidae. Exclusive
neoceratopsians are comprised of nine individual genera and three families with
their own genera species. Ranked underneath neoceratopsians is the superfamily,
Ceratopsoidea, with two genera species.
Further under ceratopsoids, is the ceratopsids under the family, Ceratopsidae. It consists of three
subfamilies, Centrosaurinae (Sin-tro-sawr-e-nay),
Ceratopsinae (Sair-ah-tops-e-nay) and
the largest neoceratopsian group of all is the ceratopsid subfamily, Chasmosaurinae. (Kaz-mo-sawr-e-nay).
 |
| Artist: Cheung-tat Chung Yinlong |
 |
| Artist: Danny Cicchetti Stenopelix |

One of the older and first successful ceratopsians
to appear was in the family, Psittacosauridae
(Sit-tah-coe-sawr-ah-day). Due to their beaks, psittacosaurids are known as the
‘parrot lizards’ and occurred in the Early Cretaceous 126-101 mya. Psittacosaurids
first arose in Asia placing all the ceratopsian groups having their ancestral
origins from Asia. Rather small, the largest in, P. mongoliensis
(mon-go-lie-in-sis) was 2m/6.5ft in total length, while P. ordosensis
(ord-o-in-sis) was 2.4m/4.6ft.
 |
| Artist: Nobu Tamura Psittacosaurids |
Psittacosaurid skulls were frill-less
and hornless, but in more derived species, bony lumps on the skull were evident
serving as a precursor to later ceratopsid horned ornaments. Juvenile fossils
suggest they were quadrupeds while transitioning into obligatory bipeds as
adults. The adult forearms could not rotate nor add lift in springing forward
giving more indication of a bipedal gait.
 |
| Psittacosaurid stomach stones |
Dentition was adapted for slicing
through plant food and due to studies of psittacosaurid teeth, the teeth
arrangement and particular wear validates that the teeth sharpened themselves.
Evidence also alludes to a seed rich diet as gastroliths have been found in
psittacosaurid fossil sites in the gut area.
 |
| Psittacosaurid fossil Note: tail feathering integuments |
There are plenty of psittacosaurid
fossils showing long quill impressions that were located on the dorsal rear and
caudal base of the animal. The quills would not have served as insulation, so
were probably for display purposes only. These preserved quills were tubular
bristle-like structures more likely with a keratinous covering and extended
through the skin almost to the vertebrae. From China’s Yixian Formation,
psittacosaurid fossils even give evidence of feathering as observed by the
studies.
 |
| Artist: Robert Nicholls Psittacosaurus sp. |
Psittacosaurid integument (body
covering) in fossil finds shows the body covered in small scales with larger
scales dispersed in irregular patterns. For camouflage purposes there were
counter shading with the top of the body exhibiting darker melanism than the
ventral portion, which was a much lighter tannish brown. This suggests a
livelihood in forests or a region with a thick canopy.
 |
| Psittacosaurid fossil family |
Due to the innumerable fossil finds,
from embryos to adults, psittacosaurids have been found in all their life
stages. From studies, hatchlings with longer forelimbs in relation to the
shorter, stubbier hind limbs would’ve been skilled crawlers. Although by five
years of age, the hind limbs underwent a massive growth squirt catching up to
and surpassing the forelimbs. By six years of age, psittacosaurids were facultative
bipeds.
 |
| Psittacosaurid fossil of six juveniles |
In the above photograph is a group of
six young psittacosaurids huddled together while being buried by a volcanic
mudflow. We know that they were alive at the time of burial because of all the
heads were raised. This huddled mass of psittacosaurids is not representative
of a nest but of gregarious behavior extending beyond the nest. Based on
histological analysis, specimen 1 was three-years-old, with specimen 2-6 being
only two-years of age. Most likely they were huddled for security due to the
fear of the oncoming onslaught that ended their lives.
 |
| Artist: Kana Hebi Leptoceratops |
 |
| Artist: Nobu Tamura Cerasinops |
 |
| Artist: Lukas Panzarin Koreaceratops |
Another primitive neoceratopsian was, Koreaceratops (Koe-ree-ah-sair-ah-tops)
that appeared towards the end of the Early Cretaceous 103 mya. It is notable
for its caudal vertebrae’s tall neural spines that were five times the height
of its back’s midpoint vertebral centra. These spines have shown up in several
other ceratopsians, such as Bagaceratops,
Montanocertops and Udanoceratops, but seems to have
independently evolved. Koreaceratops’
astragalus (talus or ankle bone) structure was also unique in that it divided
into two fossae (Fossa is a bone depression) by a prominent craniocaudal ridge
on the proximal surface. These characteristic features point to a semi-aquatic
lifestyle for swimming capabilities.
 |
| Artist: Brian Engh Aquilops bothered by a mammal |
The most primitive of neoceratopsians
are in the family, Leptoceratopsidae
that had a temporal range of 83.5-66 mya in the Late Cretaceous under ten
genera. Leptosaurids evolved in what is now Sweden and once during the Late
Cretaceous’ Campanian Stage (83.6-72.1 mya) when N. America became isolated
from northwestern Europe due to continental drift, creating endemic forms in
eastern N. America. The bulk of leptosaurids however originated from Asia and
Western N. America.
 |
| Artist: Kana Hebi Montannoceratops |
Leptoceratopsidae means ‘small
horned face’, but despite the name, leptosaurids had no horns. With species no
more than 2m/6.56 long, leptosaurid dentition set them apart from other
neoceratopsians in having massive robust jaws supporting huge bulbous teeth.
This surely was their adaptation for survival as they would never starve or be
out competed for food sources because they were able to ingest the toughest of
plants assimilated for digestion by their grinding and pulverizing teeth. Two
leptoceratopsids species in, Montanoceratops
which lived 70 mya and Leptoceratops
(Lep-toe-sair-ah-tops) occurring 66.8-66 mya both had short, deep and muscular
jaws creating a strong bite and high masticating abilities. They both possessed
claws on the distal ends of the manus (hands) and pes (feet) and also both
could move as a quadruped or bipedally.
 |
| Artist: Sergey Krasovskiy Turanoceratops |
Turanoceratops (Tu-ran-o-sair-ah-tops),
although with too numerous primitive features to be a true ceratopsid and after
numerous cladistic debates, has been put into the superfamily, Ceratopsoidea (Sair-ah-top-soi-dee-uh).
This put Turanoceratops, as basal to ceratopsids,
in representing a transition from ceratopsians to ceratopsids. At 2m/6.6ft in
length, its paired horns formed from the base of the frill pointed anteriorly
over then beyond the eyes. Turanoceratops
is the only ceratopsoid member found outside N. America. Its remains were discovered
in Uzbekistan in 90 million year old Late Cretaceous strata.
 |
| Artist: Walter Colvin Zuniceratops |
Another ceratopsoid and the first horned
ceratopsian to appear in N. America was Zuniceratops
(Zoo-nee-sair-ah-tops). The skull supported two horns above the eyes called
‘brow horns’ and a thin, broad frill that acted as a shield in protecting the
back of the head and neck. The frill had two large openings in the middle that were
covered by skin and keratinous scaling. Not exactly useful in predatory
protection, the fenestrated frill would have been lighter and was most likely
used for display.
Occurring 91 mya during the Late
Cretaceous, it was about the same size of a large cow at 3-3.5m/9.8-11.5ft in
total length. In juveniles, the dentition was single rooted, but with age in
adults, the dentition became double rooted. Whether direct or indirect through
ancestral lineage, Zuniceratops’ line
is what led to the N. American, Ceratopsidae
groups making both families a sister group.
As a sister group to ceratopsoids, the
family, Protoceratopsidae (Pro-toe-sair-ah-tops-ah-day)
occurred 85-72 mya in the Late Cretaceous. This was too late to be a direct
descendent to ceratopsids, but as a younger sister group to the older
ceratopsoid clade, they were more like an aunt to the most derived ceratopsians
in the family, Ceratopsidae.

 |
| Artist: Luis V. Rey Protoceratops |
 |
| Artist:Maspix Protoceratopsid herd in arid terrain |
 |
| Artist: Raul Martin Protoceratops/Velociraptor dueling |
 |
| Protoceratops/Velociraptor combat fossil |
A few more of protoceratopsids:
Ceratopsidae: This family group of skull horned and large frilled neoceratopsians appeared in the Late Cretaceous 83-66 mya. The horns were located on the nose and just above the eyes. With Triceratops reaching lengths of 7.9-9.0m/25.9–29.5ft and Eotriceratops topping out at 8.5m/27.9ft, ceratopsids were the largest of all ceratopsians. These dinosaurs were the Late Cretaceous version of vast herds of grazing bison on the midwestern N. American plains, except for the fact that western paleo-N. America during the Late Cretaceous wasn’t grasslands, but coastal plain marsh lands on the Western Interior Seaway with a subtropical climate and numerous small rivers and large streams.
 |
| Artist: Antonin Jury P. andrewsi |
 |
| Artist: Vlad Konstantinov a young Protoceratops |
 |
| Artist: Vlad Konstantinov P. sibiricus |
Ceratopsidae: This family group of skull horned and large frilled neoceratopsians appeared in the Late Cretaceous 83-66 mya. The horns were located on the nose and just above the eyes. With Triceratops reaching lengths of 7.9-9.0m/25.9–29.5ft and Eotriceratops topping out at 8.5m/27.9ft, ceratopsids were the largest of all ceratopsians. These dinosaurs were the Late Cretaceous version of vast herds of grazing bison on the midwestern N. American plains, except for the fact that western paleo-N. America during the Late Cretaceous wasn’t grasslands, but coastal plain marsh lands on the Western Interior Seaway with a subtropical climate and numerous small rivers and large streams.

The large frills were also ornamented
differently in each species and changed in ornamentation within each species
maturation age from juvenile to subadult to adult. Honors for the most
elaborate frill likely goes to, Kosmoceratops.
It had a row of ten small horns rimming the frill with the middle eight curving
inwards. This ceratopsid was isolated on a restricted island in the Western
Interior Seaway. The isolation diminished gene pool influxes from other parts
of N. America allowing Kosmoceratops
to evolve on its own. Once the seaway began receding southwards due to Rocky
Mountain chain orogenic uplifting, Kosmoceratops
intermingled with other diversified gene populations and lost its unique
identity.
 |
| Artist: Lukas Panzarin Kosmoceratops |
Other derived characteristics of these
two subfamilies listed as one (1) and two (2) in the cladogram below are
defined as (1) Chasmosaurinae:
enlarged rostral, presence of an interpremaxillary fossa, triangular squamosal
epoccipitals (small spikes or horns rimming the frill), rounded ventral sacrum,
and a broadly decurved ischial shaft.
(2) Centrosaurinae: premaxillary oral
margin extending below alveolar margin, jugal infratemporal flange, squamosal
much shorter parietal, six to eight parietal epoccipitals and predentary biting
surface steeply inclined laterally.


Speaking in geological timeframe terms,
ceratopsids in general evolved rapidly and simultaneously into their most
derived forms, whether genetically or through convergent evolution. As
expressed in the above graph, once centrosaurines became extinct,
chasmosaurines began evolving through convergence via centrosaurine horn characteristics
and is proven through the 68.5-67.5 million year old chasmosaurine, Regaliceratops. This 5m/16.4ft long chasmosaurine
became transitional in the skull’s horn morphologies by evolving a large
centrosaurine-like nasal horn with shorter supratemporal or ‘brow’ horns.
 |
| Artist: Luis V. Rey Einosaurus |
 |
| Artist: Vlad Konstantinov Triceratops |
The chief predator to Triceratops was Tyrannasaurus rex as evidenced through the multiple fossil finds of
T. rex gnaw marks on Triceratops
fossilized bone and actual T. rex teeth embedded in or lying near Triceratops fossils. A Montana fossil
site unearthed in 1997 showed a Triceratops
skull with cone-shaped indentations matching the tooth tips of a typically
large T. rex. In addition, on the
left side of the skull were gaps between three deep parallel scrapes which also
match the tooth spacing of T. rex which was the only large predator in
the region during the ending of the Late Cretaceous. The healed skull scars
first show signs of infection then healing, proving that the ceratopsid
survived the encounter. But whether it won in the battle or was lucky enough to
get loose and run off, we’ll never really be proven. In Triceratops/T. rex battles, we’ll go into more detail later
under, ‘Theropoda’.
Living in the Late Cretaceous 72-71 mya,
Anchiceratops (An-chi-sair-ah-tops), is
considered a transitional and intermediate ceratopsid form closely related to
both, Monoclonius (Mon-oh-clo-nee-us)
and Triceratops (Tri-sair-uh-tops). Fossil
remains have been found in terrain sediment, but in Alberta’s lower ‘Horseshoe
Canyon Formation’, where numerous fossils of it have been found is composed of
shallow marine coastal sedimentation, coal swamps, floodplains, brackish water
deposits from periodic seawater incursions into estuary channels, estuarine
point bar deposits, carbonaceous shales, mudstones and sandstone freshwater
sediment. From this, it can reasonably be interpreted that Anchiceratops was hippo-like in foraging aquatic plant material.
Another trait to allude to this conclusion is in this chasmosaurine’s skeletal
anatomy featuring a much longer pelvis, more robust forelimbs and shorter tail
than other ceratopsids.
 |
| Artist: Luis V. Rey Triceratops vs. T. rex |
 |
| Artist: Craig Dylke Anchiceratops |
Some more pictures of ceratopsids:
 |
| Credit: Phil Tippets' 'Prehistoric Beast' Monoclonius |
 |
| Artist: Julius T. Cstonyi Regaliceratops |
 |
| Artist: Nobu Tamura Centrosaurines |
 |
| Artist: Nobu Tamura Chasmosaurines |
Due to the lengthiness, Dinos will be completed with the story of theropods and birds in the next two articles of ‘Et Tunc Nulla Erat IX and X’.
Hoping Every One had a Fun Holiday Season, in Spite of Trump!
BJA
Oct 2017-Jan
2018
This comment has been removed by a blog administrator.
ReplyDeletekuşadası
ReplyDeletemilas
çeşme
bağcılar
tekirdağ
1XJQ7