Et tunc nulla erat V
(And Then There Was)
REPTILIA:
Et
Tunc Nulla Erat IV
led us through the history of amphibian evolution from very early
labyrinthodonts up to the modern or extant lissamphibians. In the attempt to
continue on in the saga of life’s further evolution, we’re going to now
introduce the reptile line that arose from diapsids slugging through extinct,
lizards turtles, amphisbaenians and marine reptiles the euryapsids, while ending
with some extant groups; but first, a better introduction to cladograms.
Cladogram is from Greek with clados meaning ‘branch’ and gramma emphasizing ‘character’. We will
be using the cladistic diagrams more with reptiles in tracing ancestral lines
instead of graphs of family trees. Cladograms do not show direct ancestral
relationships to species descendants, nor do they display evolved changes as
evolutionary trees do, but a cladogram does point to common ancestors of a
group. Cladograms can also utilize relations morphologically (et al: invertebrate/vertebrate,
prokaryote/eukaryote, skull types: synapsid/diapsid) or as a molecular unit in
DNA sequencing.

As an aid in understanding cladograms,
the first graph shows that it is the baseline as always being the ancestral
original root with speciation commencing from there. The second graph depicts
two simple forms of cladograms with speciation always arising from the base
stem. The third and fourth cladogram graph gives more detail in showing the
ancestry base’s descendants and in what order was the earliest to the latest by
numbering or naming the lines normally from oldest to younger as from left to
right or from top to bottom.

Simply put, cladograms are straight
lines with a main common ancestral line amidst other lines branching off from
it in representing different clades of the common ancestral main line.
Towards the end of the Carboniferous and
into the early Permian, atmospheric oxygen levels peaked at ~32% while
displacing higher oxygen levels into the oceans serving as sinks. This high
level created some of the largest insects with Meganeura, a dragonfly reaching lengths of 66cm/26in. That’s the
size of some sea gulls. The millipede, Arthropleura
armata reached lengths of up to 2.3m/8.5ft. Surviving into the middle of
the Permian, this arthropod was the largest invertebrate to ever walk Earth.

The higher oxygen levels benefitted
larval metabolism more so in reaching gargantuan sizes since the aquatic larvae
cannot control oxygen intake in breathing through their skin. The terrestrial
adults breathe through spiracles that can be voluntarily closed.
 |
| A model of Arthropleura |
The Permian Period lasted from ~299-252
mya. Although the beginning of the Permian was cool while ending the
Carboniferous Karoo Ice Age, the end of the Permian experienced a super mass
extinction that almost annihilated all marine and terrestrial animals due to
heated climatic change. It appears the culprit was a combination of events in the
last million years of the Permian that culminated into the wholesale slaughter.
The current Siberian basalt flood plains
~ 6,500m/21,325.5ft thick, is indicative of a million years of intensive
volcanic eruptions during the end of the Permian. During that time frame, there
was an extensive release of volcanic carbon dioxide and other obnoxious gaseous
releases that in turn warmed the atmosphere and ocean waters releasing an
immense amount of methane into the oceans that was tied up in methane hydrate reservoirs.
The methane in turn consumed free oxygen levels making ocean zones
uninhabitable for life by creating widespread anoxic/dysoxic regions, in
particular along coastlines.
Using a new method to estimate the
oxygenation of ocean water in the past, scientists found that the ratio of
thorium and uranium isotopes changed dramatically at the time of the
extinction.
Meanwhile with acidification of the
oceans due to the upper threshold release of greenhouse gases, the atmosphere
warmed so much it resulted in ocean level increases and hot climatic zones that
made it lethally too hot for life to exist along the equator and dry interior
portion within the Pangaea continent.
Today, Global Warming deniers will tell
you that warming of the atmosphere is a natural event and has occurred many
times over Earth’s history. Yes it has, but they leave it at that. What they
don’t include is its following impact and effect on life.
Fortunately, most of the earth’s Global
Warming events were small scale in the dispersion of carbon dioxide allowing
the climate and life’s adaptations to compensate and recover. Unfortunately for
life at the Permian/Triassic boundary, the change was rapid and intense killing
off most of life. Whether you agree or disagree if the current carbon dioxide
emission climatic warming is natural or manmade, it doesn’t matter, for in also
being a rapid transition, even more so in concentration than the Permian ending
witnessed, life is going to have a hard time adjusting.
The warmest NASA /NOAA global year on
record was this past 2015. It beat the previous warmest global land/oceanic
surface record, which was just the year before in 2014. To put these back-to-back records into
context, if temperatures hadn’t been rising recently, previous normal
back-to-back record high temperatures correlate to a chancy every 1500 years.
With prevailing temperature rises that is now correlating into every ten year
intervals.

Struggling to survive with this latest
event where carbon dioxide atmospheric concentrations have already surpassed
400 ppm, a sixth massive extinction is poised. Present carbon dioxide
concentrations have more than doubled the 200 ppm concentrations that current
life has adjusted to.
Now, with that over with, let’s dig into
reptiles…
The
Road to Reptiles:
Reptiliomorpha (phonetics:
Rep-til-e-o-mor-pha), otherwise sometimes listed as Anthracosauria (phonetics: An-thra-co-sor-ee-ah), is a stem clade
order deriving from a temnospondyl line. They are an ancestral lineage,
therefore more related to reptiles, birds and mammals than they are to
lissamphibians. The laying of the first terrestrial egg occurred some 340 mya
in the Mississippian Period of the Carboniferous by a reptiliomorph and most
likely one that still spent most of its days in aquatic environments, but could
definitely roam land possessing stiffening knee joints geared for a more
pronounced gait in favoring walking.
Scalation was clearly a convergent
parallel evolutionary trend from both groups of labyrinthodonts. Convergence is
morphological similarities that were acquired independently in different
species that are not related as opposed to homology. When morphologies are
homologous, it is due to two species having a common ancestor that the
morphology was inherited from.
 |
| Anthracosaurus |
In many of the lepospondyl microsaur
species, such as Trimerorhachis (phonetics:
Try-mer-or-ah-chis), overall body dermal scale patterns are not unlike
primitive reptile scalation. Trachystegos
(phonetics: tray-key-stay-ghos) of the latter Carboniferous and Saxonerpeton (phonetics:
Sax-unh-ner-pe-tawn) of the Early Permian had an extensive scale covering from
head to tail.
 |
| Saxonerpeton |
This terrestrial egg or amniote was
completely self-sufficient from desiccation in a dry environment, therefore a
huge advantage for the continuing survival of a species in arid conditions. At
the end of the Permian Period which ended the Paleozoic Era, the continents
Euramerica and Gondwana became one forming the supercontinent Pangaea. The
interior, far from coastal regions became very arid, so the steamy swamplands
of the latter Paleozoic gave way to drier conditions.
Amniotes had replaced buccal pumping for
breathing relying instead on a complex and an in-folded pair of lungs. Early
day amniotes also possessed a longer, more flexible neck devolving the skull’s
otic notch in making for a more terrestrial lifestyle.
As mentioned above, known as the ‘Great
Dying’, the Permian/Triassic extinction resulted in the largest mass extinction
ever causing ~72% of all tetrapods to go extinct. Thus ended the reign of the
amphibian-like labyrinthodont, which was still wholly dependent in laying
jelly-like egg masses in watery environs. The amphibian’s environment had gone
from widespread to isolate. During this extinction episode, even insects were
scathed losing up to 83% of all insect genera. However, this left room for a
small indiscrete animal known as a reptiliomorph to raise a legion into dominance…the
reptile…whose egg was suited for the transition. As a result, reptiles began to
rule the land, arboreal settings, air, and waters.
 |
| Reptilian amniotic egg vs. Amphibian egg |
Going from outside inwards, the amniote
egg consists of an outer shell, albumen, another protective layer called the
chorion where inside it rests the yolk sac (nutrients reservoir), allantois sac
(respiration facilitation/waste receptor) and finally within the amnion, rests
the embryo. In theriodont eutherian and metaeutherian animals extinct and
extant, the placenta affords the protection served by the shell, so in mammal
species, the chorion is the first protective membrane.
Animals such as most fish and amphibians
that lay non-amnion eggs are considered in contrast to amniotes as anamniotes.
 |
| Amniote Clad |
The exact line where reptiliomorphs
evolved from labyrinthodonts is very blurred as the reproductive units,
including eggs, fossilize very poorly. There is however fossilized evidence of
amniote reptiliomorphs occurring during the Carboniferous 340 mya. The fossil
remains are from a transitional animal containing both amphibian and reptilian
features that were laid down in very arid conditioned sediment. This animal was
the small Casineria (phonetics:
Cass-in-eh-ree-ah).
With a total length of only 15cm/5.9in, Casineria lived and laid eggs in dry
conditions hunting down the multiple invertebrates for food and possibly
finding moist spots to lay its buried eggs. Its fossil remains have only been
found in dry imposed bedrock.
 |
| Casinera |
Casineria would have
appeared as a small lizard in life complete with a scaled body and possessed on
its five toes the first known claws. Indeed, it is the first truly known fully
terrestrial tetrapod, if not the first reptilian animal.
 |
| Westlothiana |
Another early reptiliomorph is Westlothiana (phonetics:
West-lo-thee-an-ah) that occurred 338 mya. Its overall anatomy is also small at
20cm/7.9in. In the body plan there were short legs as in labyrinthodont
temnospondyls, while reptilian characteristics were unfused ankle bones and non
labyrinthodont infolding of dentin and it laid leathery amniotic eggs.
Both these animals had no otic notch and
in being small they laid small eggs. The smaller eggs experienced a much lower
volume to surface ratio. This was an advantage in developing primitive amniotic
eggs which first arose from non-amniotic terrain egg laying labyrinthodonts
that sought out moist environs such as moss covered ground or stumps that
trapped moisture.


Westlothiana is most likely
a stem clade for diadectomorphs. Diadectidae
(phonetics: Di-uh-dec-tuh-day) was a family of reptiliomorphs that although
still retained long sharp teeth protruding from the mouth had evolved from an
insectivorous diet to vegetarian. The most primitive diadectomorphans like
limnoscelids were omnivorous. Eating an insect and the plant that it was on,
over time, the latter diadectomorphs developed a plant diet until the last of
the group, the diadectids were strictly herbivores and most likely the first
tetrapod to subsist solely on plants. Diadectids were also the largest land
animals of their time in the Early Permian with the genus Diadectes (phonetics: Di-uh-dek-tes) reaching 3m/9.9ft; thus, the
measure for larger carnivores to appear.

Eggs from these trending reptiliomorph
labyrinthodonts housed non-feeding tadpoles that hatched not as tadpoles, as
they fully metamorphosed as miniature adults within the egg. Moisture from the
eggs’ ambient environment protected the eggs from desiccation through infusion.
The fetus now developed all its larval stages within the watery domain of the
egg instead of in bodies of water like the tadpole was and is still today
chained to.
About this time in the Mid-Late
Carboniferous, the reptiliomorph Gephyrostegus
(phonetics: Gee-fy-ro-steg-us) had already veered away from temnospondyls
creating its own order of Gephyrostegida.
It had tiny circular scales covering the body, large eyes and insectivorous
pointed teeth. Gephyrostegus had
fused small ankle bones into one large bone known as the astragalus; a trait
found in all later reptiles. This small creature was leading to convergent
reptilian forms, but died out during the Late Carboniferous so is more
distantly related.
What the family Gephyrostegidae offers us is not a vast amount of species, as there were only two genera, but the first real proof of amniote eggs going back to 347 mya in Visean strata of the Middle Mississippian. In a 310 million year old fossil find in the German, Westphalian formation was the well preserved remains of Gephyrostegus watsoni. The Westphalian layers are a regional part of the middle Pennsylvanian stratigraphy.
 |
| Gephyrostegus |
What the family Gephyrostegidae offers us is not a vast amount of species, as there were only two genera, but the first real proof of amniote eggs going back to 347 mya in Visean strata of the Middle Mississippian. In a 310 million year old fossil find in the German, Westphalian formation was the well preserved remains of Gephyrostegus watsoni. The Westphalian layers are a regional part of the middle Pennsylvanian stratigraphy.
 |
| Gephyrostegus skull |
Tetrapod amniotes had securely made
their presence and no longer limited to water bodies for fetal development, could
spread throughout the lands where later isolated from geological events, would
evolve into more species in evolutionary responses to ambient ecologies. Thus,
the amniote accelerated their speciation.
The amniote class, Reptilia (phonetics: Rep-till-ee-ah) has been divided into four
main subclasses based on the number of temporal fenestrae/openings positioned
posteriorly behind the eye and below or above the postorbitals as laterally on
each side of the skull. These holes served as jaw muscle attachments.
1. Anapsida ~ no openings present;
‘proto-reptiles’, turtles
2. Diapsida ~ 2 skull openings below
postorbital on each side; ‘majority of reptiles’, bird progenitors
3. Synapsida ~ 1 skull opening below
postorbital on each side; ‘mammal-like reptiles’ mammal progenitors, in which
mammals retained synapsid skulls
4. Euryapsida ~ 1 skull opening above postorbital
on each side; ‘extinct marine reptiles’ derived from diapsids losing one lower
temporal fenestra

This is a simplified view, but is an orderly scheme. No less, due to phylogeny in monophyletic and paraphyletic realms, there are much more complicated cladistics one can view and study.
Anapsids include the earliest forms of
reptiles along with all turtles. Early anapsid reptiles are sometimes referred
to as ‘parareptiles’; the term means “at the side of reptiles.” Just as
diapsids, synapsids came from the amniote line but were the first to arrive in
diverging before the diapsids’ arrival as witnessed in the cladogram below.
Synapsids include the extinct pelycosaurs and mammal-like reptiles (therapsids)
in which mammals evolved from. Mammals are the only extant synapsid group.
Diapsids include dinosaurs, crocodilians, tuataras, snakes, lizards and birds.
Even though birds no longer have any temporal fenestra, they derived from the
group phylogenetically. Euryapsids are extinct marine reptiles such as
ichthyosaurs, plesiosaurs and the lesser known nothosaurs.


Before we continue here, I’d like to
relate a side note on a parareptile. The superorder Sauropterygia (lizard flippers) includes all reptiles evolving from
land reptiles only to take to the seas as euryapsids. These creatures show up
in the middle of the Triassic to become all extinct by the end of the
Cretaceous.
 |
| Mesosaurus |
The interesting oddball we’re to
discourse a bit on here is the mesosaur, who by all intents and purposes is a
reptile but are now grouped as ‘parareptiles’. They first appear in the fossil
record 299 mya which was 57 million years before the first sauropterygian. Mesosaurs
(not to be confused with mosasaurs) were amniotes, but instead of being
anapsid, or even a diapsid derived as a euryapsid, they had a synapsid skull.
Mesosaurs had already left land to swim in the salty sea shallows or the inland
hypersaline watery environment in what is now the Mangrullo formation of
Uruguay while amphibians still ruled the land. Mesosaurs are one of the first
or very first tetrapod to return back to the seas. All other synapsids would
remain terrestrial and with a few, eventually leading to mammals.
 |
| Drawing of Mesosaurus fossil embryo |
Fossilized mesosaur embryos exhibit pachyostosis in having thicker ribs than land tetrapodal embryos. This suggests embryos in utero were ovoviviparous being born live in the water as numerous sea snakes currently do. Mesosaurs may turn out to be the most basal marine reptile, or a stem ancestor to true reptilians, or a basal and stem to both groups of parareptiles and synapsids. Or, it may simply continue to be an oddball synapsid that diverted away from all other land synapsids before mammals to live a life in the seas.
During the early stages of the Permian,
marine life took a big hit with it being rare to find any marine fossils.
Calcareous algae and calcareous sponges became the primary shallow water reef
builders. Trilobites were declining and had gone extinct by the end of the
Permian, while lobe-finned fish that had given rise to tetrapods and spiny
fishes were giving way to bony fish. Sharks maintained a stable population. On
land, fungi had finally figured out a way to consume bark 60 million years
after trees had evolved it during the Carboniferous. That is why the
Carboniferous sediment has so much coal deposits, as the tree, bark and all
were covered over. Glossopteris (an extinct order of seed ferns) came into
existence along with clubmosses, horsetails and various ferns still making
their presence as holdovers from the Carboniferous. Swamp forests began giving
way to conifer forests such as the short palm cycads and the gingko of
today.
The
Reptile:
The Early Triassic Period started off
hot with vast interior deserts and warm Polar Regions supporting lush forests
and warm shallow seas.
Thus far, we have reptiliomorphs that
had evolved from the labyrinthodont temnospondyls. This led to the
reptiliomorph sister clade, the amniote that had the ability to lay viable eggs
on land. From amniotes, the stem reptiliomorph genera Casineria and Westlothiana
as explained above, had emerged creeping ever closer to a true reptile.
Casineria, was one of the
first to have toughened feet for land equipped with claws for grasping. It was
also one of the first to develop sex as we know of it. For eggs to be protected
on land the shell formed, but sperm cannot penetrate the shell to fertilize the
egg, so internal fertilization first arose with Casineria to intercourse fertilization before the shell formed.
 |
| Paleothyris |
Paleothyris (phonetics: Pay-lee-o-thy-ris)
occurred in the mid-late Pennsylvanian of the Carboniferous from 310 to 302
mya. It most definitely was a reptile, but a very primitive one with an
anatomic reptilian body, but also supported a temnospondyli-like, though
advanced skull. It was large eyed with sharp teeth most likely hunting insects
nocturnally. As in all former labyrinthodonts it was an anapsid with no
fenestrae. The evolutionary key to this creature is that it derived from an
amniotic lineage inching ever closer to a fully reptilian trait.
 |
| Hylonomus |
From here the first true reptile evolved
in the form of the genus, Hylonomus
(phonetics: Hy-lon-o-mus). It had the typical anapsid boxed head with skull
openings only for nostrils and eyes.
Anapsida:
 |
| Anapsida Cladogram |
Hylonomus, a romeriid anapsid lived during
the Carboniferous in the mid Pennsylvanian 312 mya. It may even be older at 315
mya as fossilized footprint impressions have been attributed to this animal
after careful study of its fossilized foot bones. At 20cm/8in, it appeared as a
small lizard living and hiding in tree stumps as most fossils have been located
in fossilized club moss stumps. Hylonomus
most likely ate small invertebrates with its small sharp teeth supported by its
anapsid skull.
Surely a more ancient reptile fossil
will be found, but for as it stands, Hylonomus
is the basal ancestor to all reptiles including dinosaurs, extant crocodiles
and all mammals and birds.
 |
| Captorhinis agouti |
 |
| Captorhinids |
 |
| Late Permian bunostegos species |
Aside from still lingering argument that
turtles came directly from anapsids, there are no extant anapsids. Modern day
turtles do possess an anapsid skull, but it now appears they originally came
from a diapsid ancestral lineage. As a collage of earliest primitive reptiles,
anapsids are not a monophyletic group, but rather simply a collection of the
earliest reptiles that were distantly related. Currently, all genetic molecular
studies pinpoint turtles well within the early diapsid clade.
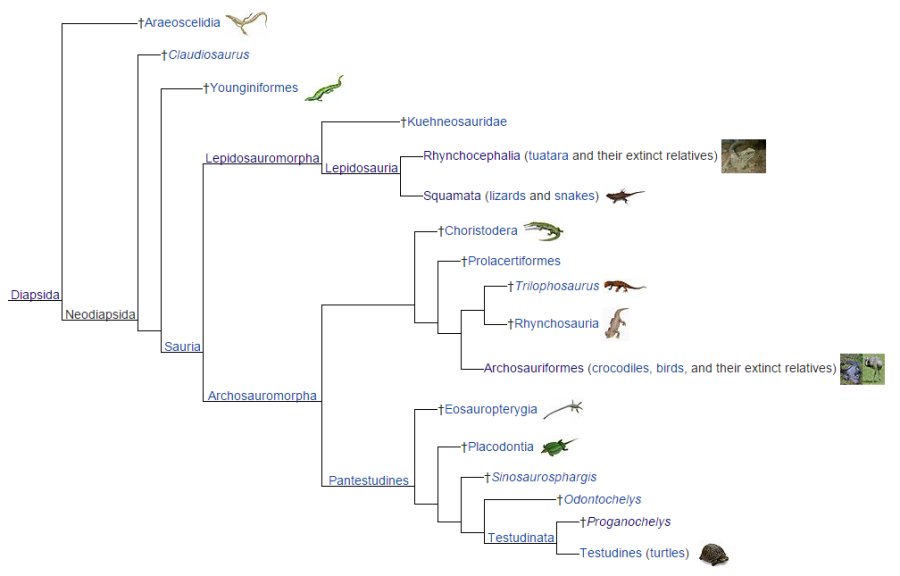
After reanalysis, Testudines (turtles, terrapins and tortoises) have been assigned to
the lepidosauromorph diapsid group that contains the lepidosaurs (snakes,
lizards and tuataras). Turtles diverged away from lepidosaurs somewhere between
200-275 mya. The turtle’s skull began reverting back from a diapsid to an anapsid
skull as the lineage began losing teeth replacing the dental ware with a
chitinous beak.
Synapsida:


Synapsids had already diverged from the
amniote tree during the Late Pennsylvanian 308 mya devolving away from true
reptile morphology. The divergence formed the Synapsida line that eventually led to mammals. The Sauropsida line would eventually lead to
diapsids which includes crocodiles, pterosaurs, marine reptiles, turtles,
lizards/snakes and dinosaurs/birds.
The undisputed oldest synapsid fossils
are Echinerpeton intermedium (phonetics:
Eh-she-nair-pah-tawn inter-med-e-um) and Archaeothyris
(phonetics: Are-kay-o-thy-ris) from 308-306 mya respectively. Echinerpeton, though only the size of a
small lizard, just might be the base ancestor to the later much larger synapsid
sail-backed dimetrodons that reached up to 4.6 m/15ft. Echinerpeton had articulated vertebrae with high neural spines
supporting a dorsal webbed sail.
 |
| Echinerpeton intermedium |
Synapsids may even have an earlier
origin in the ~314 mya fossil remains of Protoclepcydrops
(phonetics: Proto-klep-sigh-drops). Unfortunately the single skeletal remains
are fragmentary, but the disarticulated remains do allude to a synapsid skull
type. Early synapsids begin digressing away from eventual true reptile
evolution leading eventually to mammals.
The synapsid line led to pelycosaurs
including the dimetrodons that formed the basal ancestry to therapsids
(mammal-like reptiles) that eventually gave rise to mammals. For this treatise,
we’re only going to mention a few facts about pelycosaurs where more in depth
discussion will be given in a future ‘Et Tunc’ series concerning synapsids to the
rise of mammals.
Pelycosaurs are one of the earliest most
primitive of synapsids. First appearing in the Upper Pennsylvanian ending,
pelycosaurs became extinct by the end of the Permian.
 |
| Bone growth types: A.low vascular B. highly vascular C. fibrolamellar |
Warm blooded animals (endothermic/homoeothermic)
like birds and human grow at a faster rate than cold blooded (ectothermic)
animals do. With this fast growth rate process found only in homoeothermic
animals is a bone growth pattern termed fibrolamellar bone (FLB). The early pelycosaur,
Ophiacodon (phonetics:
O-fee-ah-co-don) also included FLB in its physiology. One of the greatest
opportunities in evolutionary devices is the regulation of internal warm
bloodedness. Although Ophiacodon was
not directly related to mammals and even more remotely related to birds, it had
accomplished at least some form of thermoregulation nearly 300 mya. Living
296.4-279.5 mya, it also had the largest skull of any contemporary animal of
its day reaching lengths of 50cm/20in.
 |
| Compare head sizes Ophiacodon in middle; Cotylorhynchus in back |
In direct contrast to Ophiacodon’s large skull was the
pelycosaur caseid, Cotylorhynchus’s
(phonetics: Co-tee-lor-reen-cuss) minute skull. With a barrel shaped and
heavily set body at 6m/19.7ft in length while weighing in at ~1 ton, the head
was only around 87.5cm/34.5in. The nasal openings and cavities were also large
for the skull, perhaps to take in more oxygen or functioned as a higher surface
area to moisture. Nonetheless, its girth allowed for larger organs in more
functional digestion of plant material. Being the largest herbivore during the
Permian, with the massive body used as a weight force, the front legs were used
for digging up roots. Regardless of the animal’s small head, its total size
also acted as a predator deterrent as from 279.5-272 mya it was quite common in
many Permian environments.
A miniature Cotylorhynchus was Casea
(phonetics: Cah-say-ah) at 1.2m/4ft. Its ribcage was highly expanded to make
way for a large gut to properly digest plants like horsetails and ferns. It
also was successful with fossils being found from Texas to France.
 |
| Varanops |
Varanops (phonetics:
Va-ran-ops) has the distinction to being one of the very last pelycosaurs to
die off becoming extinct at the end of the Permian. Perhaps this was due at
least in part to competition against an established rising number of therapsids
during that time.
 |
| Dimetrodon |
 |
| Edaphosaur |
Diapsida:
Through Sauropsida (phonetics: Saur-op-see-da) lineage, two major
stem-based clades diverged forming the Eureptilia
(phonetices: U-rep-til-e-ah) and the Anapsida/‘Parareptilia’. Out of Eureptila, the Diapsida clade evolved.
 |
| Eureptilia Clade |
Around the Carboniferous/Permian boundary, two diapsid groups diverged known as the Neodiapsida (phonetics: Nee-o-die-app-sah-duh) and Araeoscelilidia (phonetics: Ah-ray-oss-kell-ah-lee-dee-ah). One early diapsid araeoscelilid was Petrolacaosaurus (phonetics: Pet-row-lak-oh-sor-us).
 |
| Petrolacosaurus |
Petrolacosaurus appears in the
Late Carboniferous 302 mya and as a direct line had died out by 275 mya.
Superficially similar to modern day lizards, this rather small extinct reptile
at 40cm/16in is the first known amniote diapsid and possessed canine-like
secondary teeth as latter forms of therapsids and mammals will. It even
possessed a heart template as all modern mammals do, including humankind. Even
though definitive evidence is not in yet, petrolacosaurids just might be a common
ancestor to both diapsids and synapsids as a basal diapsid amniote. The
well-formed two temporal fenestrae in its phylogeny might have had one deleted
as the turtle line had deleted both. In evaluating its teeth, most likely Petrolacosaurus was an insectivore.
 |
| Neodiapsid Clade |
Another araeoscelilid was Spinoaequalis (phonetics: Spy-no-aye-kwal-iss).
Occurring 300 mya it had longer and stronger hind limbs. Its symmetrical spine
supported a laterally flattened tail used for propulsion in water. Spinoaequalis was at home on land as
well as water using its tail for propulsion. This reptile was semi aquatic and
may have not only frequented freshwater, but transitioned to marine as well for
some of its fossil finds are in the same sediment as some well-preserved fish
fossils.
 |
| Spinoaequalis |
Around 260 mya, Neodiapsida further diverged into the 60cm/2ft Claudiosaurus (phonetics: Claw-dee-o-saur-us) and Sauria (phonetics:
Sawr-ee-ah). One of the earliest neodiapsids forming the basal diapsid group,
claudiosaurids had a poorly developed sternum, more cartilage than bone, a
relative long body/neck and swam marine shorelines frequenting rocks above the
surface to sunbathe living much like marine iguanas today in sunbathing on land
and swimming in sinusoidal undulating fashion with legs tucked to the sides.
Although it had developed limbs, but with webbing, due to the amount of
cartilage in its osteology and poorly developed sternum, it most likely rarely
frequented land.


With the elongate body/neck, same
paddle-shaped distal limbs, lack of a lower skull temporal bar and transvers
flange of the pterygoid, a closed condition of the palate and reduced
suborbital fenestrae, claudiosaurs are seriously considered as a basal ancestor
to nothosaurids and plesiosaurids. Other than that the claudiosaurid line
became extinct towards the end of the Permian 253 mya.
 |
| Sauria (Archelosauria branch) cladogram |
The other neodiapsid divergence, the Sauria clade was once relegated only to
the lizard line, but since the introduction of molecular genetics, it has been
broadened to include the most recent common ancestors to lepidosaurs and
archosaurs. So, all lizards, snakes, crocodilians, birds, tuataras and numerous
extinct animals such as pterosaurs, marine reptiles and dinosaurs are saurians.
This new rearrangement has aided in distinguishing stem saurians such as in the
synapomorphy characters of cephalad (pertaining to anterior or posterior ends)
regions, the trunk, pectoral, pelvic and limb sectors.
 |
| Sauria (Ankylopoda branch) cladogram |

The saurian clade represents the divergence between lizards and crocodiles while the clade itself is subdivided further into Lepidosauromorpha (phonetics: La-peed-o-sor-o-mor-pha) meaning “closer to snakes than birds” and Archosauromorpha (phonetics: Ar-ko-sor-o-mor-pha) meaning “closer to birds than snakes.” An early period saurian group was in the genus, Coelurosauravus (phonetics: See-lore-oh-sor-ay-vuss). This genus was one of the first lizard groups to take to the air ~260.5 mya. Coelurosaurvus will be discussed later under lizard evolution.
 |
| Coelurosaurus skeletal |
Teeth type plays important roles in
evolutionary niches. Thecodontia teeth once even played a role in taxonomic
grouping, although it now is obsolete. Thecodont means ‘socket tooth’ and
represents animals with teeth socketed through the gums into the jawbone.
 |
| Tooth Implantaions: A. acrodont B. pleurodont C. subthecodont D. thecodont |
Pleurodonty (pleurodont ~ side tooth)
exhibits teeth ankylosed (fused) by their sides to the jaws inner surface.
Acrodonty (acrodont ~ summit tooth) exhibits teeth mounted in a slight
depression atop the gums and are not socketed with roots. Sphenodonts,
pleurosaurs and certain extant lizards will utilize this teeth arrangement.
 |
| Youngina red possible nascent antorbital fenestra |
Youngina (phonetics:
Yun-gin-ah) is a basal lepidosauromorph that occurred during the Late Permian.
This lizard-like reptile was 1.47m/4.82ft long and with its conical teeth, was
most likely an insectivore, but also carnivorous to small vertebrates. The
photo below is a Youngina skull. The
red highlights a possible nascent antorbital fenestra.
From here, the diapsid clade Archosauromorpha came about 275mya. The skull opening between the nostrils and
eye sockets is known as the antorbital fenestra. In the archosauromorph family,
Protorothyrididae (phonetics:
Pro-tor-o-thy-rid-a-day), the genus Protorosaurus
(phonetics: Pro-tor-o-sor-us), which means ‘first lizard’ is the earliest known
archosauromorph thus far. Archosauromorphs would lead to Archosauriformes (Phonetics: Ar-ko-sor-e-forms) that eventually led
to true archosaurs that would further lead to the only extant archosaur lines,
which is of course the crocodilian and avian groups.
Most of these transformations from Archosauromorpha to Archosauriformes and Lepidosauromorpha
to Lepidosauriformes occurred right
before or just after the Permian/Triassic extinction. The Archosauriformes and Lepidosauriformes
survived, the archosauromorphs and lepidosauromorphs didn’t.
The
Path to Turtles:
Turtles were one of the first diapsid
archosauromorphs to branch off the archosaur line through the Pantestudines (phonetics: Pan-tess-tu-dee-nees)
group, via the lepidosauromorph infraclass. While in the process, turtles
reverted back to an anapsid skull; so in the proceeding course, they did not
evolve directly from the early reptilian anapsids.
 |
| Pantestudines Clade |
In turtle genomics there are diverse
chromosome numbers and chromosomal rearrangements. Before developing a shell
that is fused to the ribs, turtles first had to have broadened ribs; this is
evident in the Pantestudines clade.
 |
| Placodus |
The earliest placodonts looked much like
a modern day iguana, only much larger. Placodus
(Phonetics: Plaque-o-dus) at 1-2m/3.3-6.6ft consisted chiefly upon a durophagus
diet such as shellfish and animals with hard exoskeletons like crabs. Placodus possessed chisel-like front
teeth and broad flattened back teeth for crushing prey. It must have been a
shore animal frequenting shallow waters to obtain its shell-fished prey.
Although the short limbs ended in claws, in between the digits webbing was
present and with a flattened tail, these features aided in propelling the
animal in water.
Latter placodonts such as Cyamodus (phonetics: Sigh-am-o-dus) and Henodus (phonetics: Hen-o-dus), due to
convergent evolution looked superficially like a turtle with a carapace and
plastron. Most likely this armory was developed as protection due to an
increase in larger marine reptile predators entering placodont shallow coastal
water environments. Cyamodus even
possessed a second shell protecting the rear pelvis area. A second advantage to
the shells was their increased weight that was just below the level of neutral
buoyancy allowing the animal to paddle in reaching the shallow bottoms where
the shelled bivalves were.
 |
| Placodont Henodus |
 |
| Eunotosaurus |
Occurring 265.8-251 mya, Eunotosaurus (phonetics:
Eu-no-toe-sore-us) was an early Pantestudines
still retaining the diapsid skull. This reptile had a broad body formed by nine
pairs of broadened ribs. The upper surface of the ribs was concave giving the
body a rounded broad shape. Most of the ribs were fused to the vertebrae.
Sharpey’s fibers aid in anchoring muscle to bone in most animals. On the ribs’
anterior sides, Eunotosaurus lacked
Sharpey’s fibers suggesting the animal lacked intercostal muscles. Turtles also
lack intercostal muscles.

Another Pantestudines arising 240 mya during the Middle Triassic, was Pappochelys (phonetics: Pa-po-sha-lis). This animal had exhibited pachyostosis (expanded broad ribs/vertebrae) and had gastralia, which are dermal bones found in the ventral body walls. The gastralium appears to be a precursor to a shell. It was still a diapsid in possessing two temporal fenestrae skull holes.

Another Pantestudines arising 240 mya during the Middle Triassic, was Pappochelys (phonetics: Pa-po-sha-lis). This animal had exhibited pachyostosis (expanded broad ribs/vertebrae) and had gastralia, which are dermal bones found in the ventral body walls. The gastralium appears to be a precursor to a shell. It was still a diapsid in possessing two temporal fenestrae skull holes.
 |
| Pappochelys |
 |
| Odontochelys |
With the acromion advent, turtles moved their shoulder blades and pelvis (hips) underneath their ribs. The monophyletic sister clade to Odontochelys is Testudinata (phonetics: Tess-tu-dee-nah-ta) forming the two modern turtle groups in the genus, Proganochelys (Phonetics: Pro-gan-oh-kell-ees) and the order of extant turtles, Testudines (Phonetics: Tess-tu-dee-nees).
 |
| Add caption |

In the embryonic stage, the chicken,
mouse and turtle are all similar resulting from a common ancestor. But, as the
embryo develops with the ribs extending down the embryo’s flanks surrounded by
the myotome (muscle plate), where the chicken and mouse’s ribs, scalpulae
muscle plate stay in place as the embryo matures, hatches and grows into an
adult, the turtle embryo folds the myotome forward while the ribs, as shorter,
never reach down into the flanks.
 |
| Desmayochelys |
The oldest known turtle to show the
modern features of extant turtles was in the sea turtle Desmayochelys (Phonetics: Des-may-o-kell-ees) occurring 120 mya in
the Mid-Cretaceous. However, this ancient turtle evolved independently from
modern sea turtle lineage. This suggests that there were multiple instances of
land dwelling turtles evolving into sea turtles.
Testudines is further
divided into two suborders, Cryptodira
phonetics: Crypt-o-deer-ah) and Pleurodira
(phonetics: Pleur-o-deer-ah). All extant turtles have eight neck vertebrae, but
in Pleurodira the cervical vertebrae
are spool-shaped in cross-section with biconvex centra on one or more of the
cervicals. Pleurodires include the side neck turtles that withdraw the head
into the shell by bending the neck into a horizontal plane. Cryptodires include
all other turtles, terrapins and tortoises that pull their heads straight back
into the shell.
Two really large extinct turtles were the pleurodires, Carbonemys (Phonetics: Car-bon-em-iss) and Protostega (phonetics: Pro-toe-stay-gah). Carbonemys existed 60 mya in what is now Columbia. This terrestrial freshwater turtle’s shell reached a length of 1.72m/5ft 8in. Though not definitive, it was a carnivore and could easily consume a small crocodile with super strong jaws.
 |
| Carbonemys |
Two really large extinct turtles were the pleurodires, Carbonemys (Phonetics: Car-bon-em-iss) and Protostega (phonetics: Pro-toe-stay-gah). Carbonemys existed 60 mya in what is now Columbia. This terrestrial freshwater turtle’s shell reached a length of 1.72m/5ft 8in. Though not definitive, it was a carnivore and could easily consume a small crocodile with super strong jaws.

 |
| The pig-nosed Arvinachelys |
Just named Arvinachelys (phonetics: R-vin-ack-ell-iss), a recent 2015 fossil
find is a .61m /2ft long turtle that once roamed the bayous and lazy rivers of
present day Utah ~76 mya in the Late Cretaceous. It’s pig-like nostril snout
makes it a very peculiar turtle indeed baffling scientists on what exactly was
its function.
 |
| Puentemys |
In 2012, the fossil remains in what is
now Columbia, Puentemys (phonetics:
Pwen-twem-iss) was a turtle with a carapace that extended 1.5m/5ft in diameter.
In addition, the shell was almost a perfect circle. Puentemys lived 60 mya along with enormous snakes and crocodilians.
The carapace size would have made the turtle much less susceptible to being on
the larger reptiles’ menu.
 |
| The last embrace |
Please note the above photo. One turtle
fossil that is more important than the turtle itself is of Allaeochelys (phonetics: Al-ah-ee-ock-ell-iss). The male, smaller
than the female were both caught in a loving embrace that’s forever preserved
in fossilization. The pair died mating, with scientists surmising that the
couple started the reproductive ritual near the surface of a lake and once
embraced sunk to the bottom where a layer of poisonous gases existed perishing
the couple. In the German Messel Pit, not only one fossil has been found of
this turtle mating, but no less than nine have been found, although in varying
time zones during the Eocene 57-36 mya. Perhaps as suggested, a more befitting
name would be ‘Coitus interruptus’.
The
Path to Sphenodonts/Lizards/Snakes/Amphisbaenians:
As mentioned above, during diapsid
appearances, the mainline was the Archosauromorpha
infraclass which first appeared ~255 mya. A few million years later diverging
away from archosauromorphs ~ 247.5 mya was lepidosauromorphs, in which
comprised all diapsids closer to lizards than to archosaurs. Lepidosauriformes, as predecessors to
the lepidosaur extant squamate line of lizards, snakes, amphisbaenians and the
sphenodont tuataras were already lizard-like in physical appearance.
Taking up where we left off in
discussing lepidosauromorphs under ‘Diapsids’, lepidosauromorphs had produced
the Lepidosauriformes clade. This
group is almost there as a lizard. The supratemporal of the skull is reduced
while migrating backwards replaced in its original position by the squamosal
that rims the upper temporal fenestra as witnessed in true lizards.
 |
| Coelurosauravus |
In the Late Permian ~ 260.5-251mya and
in the Late Triassic ~210 mya, two lepidosauriforms emerged in the
aforementioned Coelurosauravus (phonetics:
See-lor-o-sor-ay-vuss) and the Kuehneosaurus
(phonetics: Keen-e-o-sor-us) genera respectively. Both possessed longer tails
as opposed to their predecessors and had a streamlined rigid body. They also
had longer claws for grasping onto the barks of trees in an arboreal lifestyle.
The more ancient Coelurosauravus (40cm/16in) had a skull crest where Kuehneosaurus
(72cm/2.3ft) didn’t, but both had thin hollow skeletal structures for
aerodynamic flight. The winged assembly of Coelurosauravus
was held by newly evolved dermal bone extensions ending in posterior
pseudo-ribs supporting a skinned membranous web known as a patagium. In the
case of Kuehneosaurus, the ribs had
been thinned and extended. These were one of the first vertebrates to take to
the air.
Coelurosauravus was a true
glider lofting from branch to branch or from tree to tree much like a flying
squirrel today does, where Kuehneosaurus
most likely parachuted instead of gliding to get from higher limb to lower limb
or from tree to ground. This wasn’t a slow parachute descent. According to 2008
aerodynamic studies by K. Stein, descending at a 45° angle, speeds could’ve
easily been reached up to 10-12 m/33-39.6ft per second.
Derived as a niece so to speak from a
sister group of Coelurosauravus, but
from a sister of Kuehneosaurus was
the genus, Icarosaurus (phonetics:
Ick-ah-ro-sor-us). This little gliding reptile, no more than 10cm/4in long, in
addition to membranous skin webbing down the sides of its body, also possessed
smaller webs along the backside of the hind legs.
Icarosaurus first appeared
228 mya. Rather than developing its long rib wings in the embryonic egg, it
most likely developed them after it hatched and matured. Its pedal proportions
of the feet were similar to Coelurosauravus,
but more in line of terrestrial lizards in every other aspect.
Sharovipteryx (phonetics: Shar-ov-ip-ter-ix) occurring 225 mya had increased the Icarosaurus’ hind patagium to foot and leg wings, however there were no front limbed patagia (plural for patagium) freeing the forelegs. With large hind limbs making the much smaller forelimbs appear diminutive in comparison, Sharovipteryx most likely was not a quadruped, but hopped around on its hind limbs when not gliding during mobility.
Xianglong (phonetics: Zang-long) is, in the fossil record, the last of these gliding reptiles occurring in the Early Cretaceous. Besides the torso extended membranes, it also had two small neck membranes used as rudders.
 |
| Icarosaurus |
Sharovipteryx (phonetics: Shar-ov-ip-ter-ix) occurring 225 mya had increased the Icarosaurus’ hind patagium to foot and leg wings, however there were no front limbed patagia (plural for patagium) freeing the forelegs. With large hind limbs making the much smaller forelimbs appear diminutive in comparison, Sharovipteryx most likely was not a quadruped, but hopped around on its hind limbs when not gliding during mobility.
 |
| Sharovipteryx |
Xianglong (phonetics: Zang-long) is, in the fossil record, the last of these gliding reptiles occurring in the Early Cretaceous. Besides the torso extended membranes, it also had two small neck membranes used as rudders.
 |
| Xianglong |
In the Late Permian 255 mya one of the
first proto-squamate appears in Lacertulus
(phonetics: Lah-sur-tull-us). It had a wider premaxilla, vomers and palatines
than lepidosaurs preceding it and has been described as a facultative bipedal
animal with longer hind limbs.
Another basal lepidosauriform was Santaisaurus (phonetics:
San-tah-ee-sor-us). Occurring during the Early Triassic, it differed in having
small sub pleurodont teeth; an augury to fully developed sphenodont teeth.
Thus, begins the Sphenodontia (honetics:
Sfen-o-don-chee-ah) order as an offshoot away from the evolutionary line of
their sister group, the squamates (lizards). The only remaining extant
sphenodont is the tuatara of New Zealand with two species.
 |
| Basal proto-sphenodont |
Splitting 220 mya in the Triassic,
proto-sphenodonts as the likes of Marmoretta
(Phonetics: Mar-mo-ret-tah) began diverging from lepidosaurs in anatomically
having distinct characteristics while superficially resembling lizards.
One of the earliest sphenodonts was in
the basal genus, Gephyrosaurus (phonetics:
Jeff-er-o-sor-us) appearing in the Early Jurassic then becoming extinct.
Synapomorphies (unique shared
characteristics) among sphenodonts are located in the mouth such as an enlarged
palatine tooth row with acrodont dentition and a posterior extension of the
dentary. Sphenodonts for the most part also displayed some form of propalinal
(forward and backward mastification) jaw action.
The five taxa that sphenodonts are
divided in are the aquatic pleurosaurs and sapheosaurs, the specialized
scissor-like teeth carnivorous clevosaurs, the Mid-Late Cretaceous highly
specialized eilenodontines and the sphenodontines which are solely represented
by the two existing tuatara species.
 |
| Pleurosaurus goldfussi |
Down through the Triassic and into the
Jurassic, there were various proto-squamates leading ever closer to true
lizards until the Early Cretaceous when one of the last proto-squamates in the
genus, Meyasaurus (phonetics:
May-ah-sor-us) morphologically appeared more lizard than not.
 |
| Astralago-calcaneum |
Meyasaurus had typical lizard traits such as the scapulocoracoid pectoral girdle arrangement and a pelvis opening called the thyroid fenestra. It also had tail vertebrae as shallow keels representing possible caudal autonomy, foot bone metacarpals and phalanges capped by cartilage, heavy ossification of limb bones and the specialized lizard foot and ankle structure of the fused astralago-calcaneun. The primary difference is that the teeth of Meyasaurus were not homodont, but were heterodont in possessing anterior small pegs with posterior bicuspids.
 |
| Sphenodont clad |
Lacertilia: Most extant lizard
groupings had their beginnings in the Cretaceous. Today’s lizards are arranged
into five infraorders; they are:
1. Iguania (among others iguanas,
crotaphytids, agamids, chameleons)
2. Gekkota (including geckos, legless
lizards, blind lizards)
3.
Scincomorpha (including among others skinks, whiptails, cordylids)
4.
Diploglossa (anguids,
anniellids, xenosaurids)
5.
Platynota (monitors,
helodermatids, mosasaurs ~ extinct)
Proto-lizards with essential lizard
characteristics branched out in forming the various lizard groups. From the
late Jurassic ~155 mya, Euposaurus (phonetics:
U-poe-sor-us) with shared sister taxa traits of skull features appears to be
the basal most to Iguania, Gekkota, Scincomorpha and Diploglossa.
On the basis of frontal morphology and limb proportions, Euposaurus nested as ancestral to Scandensia (Phonetics: Scan-den-see-ah) and Liushusaurus (Phonetics: Lee-oo-shu-sor-us).
Found in Early Cretaceous rock layers
from ~130 mya, Scandensia possessed
trenchant claws in leading an arboreal life. But this squamate came down from
the trees giving rise to the basal line of Liushusaurus
that is basal to the suborder, Scleroglossa
(Scl-ro-gloss-ah) which includes iguanas, geckos, skinks and varanoids. It also
includes the new clade, Bifurcata
(Phonetics: By-fur-caw-tah) which proposes Iguania
as a sister taxon to anguimorphs. Due to the latest phylogenetic analysis on
molecular data concerning extant squamates, Bifurcata
was instituted to make Scleroglossa
valid in distinguishing the bifurcated tongued lizards from the iguanian
muscular full tongue used for lingual capturing of food.
 |
| Yabeinosaurus fossil |
The split between Iguania and lizards contained in Scleroglossa is represented in Yabeinosaurus
(phonetics: Yah-be-in-o-sor-us) as one of the earliest splits within lizard
evolution. A Yabeinosaurus fossil
found in 2011 with fifteen developed embryos is the oldest fossil of a
live-bearing lizard. In addition, physiologically, Yabeinosaurus was trending towards a varanid body shape.
 |
| Estesia skull |
Estesia (Phonetics: A-tees-e-ah) showing up in the Late Cretaceous is basal to varanus and ancestral to Heloderma, the Gila monster genus. Estesia’s dentition alludes to the fact that it was venomous. The teeth were all sharp and recurved as in current varanoids.
 |
| Estesia drawings |
There is one varanid lizard from the
Pleistocene that went extinct just 50,000 years ago; it was Megalania (phonetics:
Meg-ah-lane-ee-ah). From what is now Australia, this is the largest lizard
known reaching a body length of 5.5m/18ft and weighing 575kg/1,268lbs. With the
tail it could’ve reached a length of 7m/23ft.
 |
| Megalania |
 |
| Megalania |
 |
| Mosasauroidea clade |
Mosasaurs are a suborder of lizard going
back to the seas while becoming an apex predator in being as dominant in the
waters as Tyrannosaurus was on land (Sea rex vs. T rex).
Roaming the shallows and ocean depths
for any sized meal, mosasaur species traveled the world’s open oceans and
inland seas. Nonetheless, mosasaurs did evolve from terrestrial origins
replacing feet and toes with flippers.
 |
| Mosasaur after a shark |
Mosasaurs breathed air so had to surface
to breathe periodically and were efficient rapid swimmers possessing a strong
rudder designed broad tail for propulsion with limbs as flippers to steer and
cut through water. Mosasaur limbed flippers have the same skeletal structure as
their terrestrial lizard ancestry, but instead of feet and toes, membranous
webbing overtook the whole foot as mosasaurs evolved encasing the foot skeletal
elements. These hydropedal limbs evolved at least twice independently in
mosasaur species.
 |
| Semi-aqautic Dallasaurus skeleton |
 |
| Mosasaur skeletons |
Occurring in the Middle Creataceous,
100-95 mya, Aigialosaurus (Phonetics:
Eye-gee-ah-lo-sor-us) lived along the Cretaceous river/lake shores, river
deltas and coast lines. It most likely ate aquatic animals living along shores.
Aigialosaurus was 1.4m/4.5ft
long and was a slender animal. It is intermediate to varanoids in having
varanoid terrestrial limbs. Thus far, being the basal most representative of
the Mosaurinae subgroup, it did have
toes with claws, but also as representative to mosasaurs, possessed mosasaurid
characteristics of fused frontal bones, a developed hinge joint between the angular
and splenial bones, a circular configuration of the quadrate bone and a
reduction of the transverse processes and zygapophyses which are one of the two
paired processes of a vertebra that interlock it with the adjacent vertebrae.
 |
| Aigialosaurus |
Appearing a few million years later in
the fossil record was Dallasaurus
(Phonetics: Dal-lah-sor-us) occurring 92 mya in the Middle Cretaceous. It is
the intermediate between Aigialosaurus
and mosasaurs in the Mosaurinae
subgroup.
 |
| Dallasaurus |
Dallasaurus’ posterior
maxillary teeth were varanoid in being strongly recurved. Along with moderately
webbed feet, this lizard was semiaquatic frequenting the waters along
shorelines to appease its piscivorous diet. Dallasaurus
also had the distinctive mosasaur fused haemal arches, a humeral postglenoid
process and an elongate atlas synopophysis to name a few.
I’m attempting to be just wise enough in
mentioning here, Coniasaurus
(Phonetics: Ko-nee-ah-sor-us) that appeared in the fossil record 97-84 mya.
Although there is still debate due to the incomplete fossil finds of two
species, phylogeny analysis and cladistics suggest that Coniasaurus is a sister group to an Aigialosaurus and mosasaur clad.
 |
| Mosasaurus |
Mosasaur evolution experienced a rapid diversification and global distribution during the Late Cretaceous. To break it down in a generalized form:
100-94 mya ~ Mosasauroidea; Aigialosaurus
92-90 mya ~ Mosasaurinae; Dallasaurus and other mosasaur
precursors
89-85.8 mya ~ first wave of primitive
but true Mosasaurs; Clidastes,
Platecarpus
85.8- 83.5 mya ~ much larger/distributed
worldwide Mosasaurs; Tylosaurus
83.5-71.3 mya ~ second wave of
Mosasaurs; Mosasaurus, Globidens,
Halisaurus
71.3-65.4 mya ~ greatest diversity even
invading freshwater; Pannoniasaurus
65.4 mya ~ all mosasaurs become extinct
Mosasaurs were the first vertebrate
animal group to populate the entire world. The map below, adapted from
paleontologist S. Suzuki’s original work (1985) shows the Late Cretaceous
distribution of mosasaurs 84-66 mya. The dashed lines represent the extent of
epicontinental shallow sea encroachment during the time period.
 |
| Mosasaur global fossil sites |
The reason for mosasaur extensive
migrations is most likely due to food competition and in seeking new niches.
During this period sharks were becoming abundant [such as Cretoxyrhina (Phonetics: Kreh-tox-see-rye-nah), a 7.6m/25ft long
prehistoric shark] and would eventually rule the seas after the K-T extinction.
With a flexible skull and a head contributing to anywhere from 10%-14% of total body length, from molluscs, fish to large euryapsids and sharks, mosasaurs ate anything they chanced upon.
There were also freshwater mosasaurs
towards the end of the Cretaceous and just like in the mass ocean migrations,
most likely it was due to predator competition and pressures from lack of prey
that induced certain mosasaurs to move upstream from river deltas further
inland.
With a flexible skull and a head contributing to anywhere from 10%-14% of total body length, from molluscs, fish to large euryapsids and sharks, mosasaurs ate anything they chanced upon.
 |
| Tylosaurus keel scaled skin fossil |
 |
| Tylosaurus mosausaur |
A Plioplatecarpus
(Phonetics: Ply-o-plat-ee-car-pus) unnamed species was found in Alberta and
Saskatchewan, Canada in 85-3-83.5 mya overbank deltaic deposits with no
evidence of opened marine environments letting out into the ocean. This
evidence confers that Plioplatecarpus
had indeed invaded estuarine and river environments. The older genus, Platecarpus (Phonetics: Plat-ee-car-pus)
had evolved into the genus, Plioplatecarpus.
From shallow coastal seas, Plioplatecarpus
moved inland into rivers.
Several mosasaur species of juveniles
and adults were found far from the seas in inland rivers deposits. The
Hungarian finds show the animal had webbed limbs with terrestrial lizard-like
feet, so apparently was in a state of evolutionary reversion back to claw-toed
limbs. This mosasaur species was named, Pannoniasaurus
inexpectatus (Phonetics: Pan-nown-nee-ah-sor-us N-ex-pec-tah-tus).
 |
| Pannoniasaurus |
Pannoniasaurus lived between
85-3-83.5 mya surviving in a freshwater river system composed of an island
chain that sat between the African and Eurasian landmasses in what was once the
Tethys Ocean. With a size of 6m/19.7ft and a mouth set full of sharp teeth, it was
the rivers’ apex predator even though prehistoric alligators shared the rivers
it inhabited.
Still maintaining mosasaurid skull
characteristics, Pannoniasaurus
further had flattened the skull much like a crocodile’s which aided in ambush
strategies on aquatic and bank frequenting animals.
 |
| The mosasaurid Tylosauarus |
Lizards, as the case with mosasaurs were
and are one of the most successful groups radiating outwards into differing
species with today exhibiting over 3,000 species. One of the main components
that added to this success was cranial kinesis allowing for skull element
flexibility.
 |
| Lizard cranial kinesis |
Cranial kinesis deals with skull
motility and is the presence of moveable joints within the cranium. All
vertebrates have some form of movable skull parts whether it’s simply the lower
jaw joint only or the flexible unhinging of skull elements.
Mesokinesis, involving the frontal and
parietal bones, experiences more jointing in the rostral region of the cranium,
while metakinesis is experienced jointing between the dermatocranium and
occipital segment. Hypokinesis is a decreased contraction within cranium parts
where jointed.
Streptostyly is defined as a rotation of
the quadrate at its dorsal articulation against the squamosal and/or
supratemporal bones. The axis of rotation is transversely oriented, so that the
quadrate swings through an anteroposterior arc. Like other forms of cranial
kinesis, streptostyly involves quantifiable movement of cranial elements applying
some kind of force against each other, whether it be tension, compression or
friction. Streptostyly is the fore-aft movement of the quadrate about the otic
joint (quadratosquamosal joint), although transverse movements may also be
possible.
Amphikinesis is the occurrence of
mesokinesis and metakinesis in a coupled and coordinated manner. For any type
of kinesis to occur there must be neurokinesis addressing movement between the
braincase and palate at the basipterygopterygoid joint in the vicinity of the
sphenoid bone.
Cranial kinesis characterizes distinctive
skull movement features. Kinesis features were slowly adapted improving the
modes of capturing and holding prey prior to swallowing. By the time snakes
evolved, cranial kinesis, along with neurokinesis were already well evolved and
genetically inherited.


Serpentine: The big battle
over snake evolution on whether it evolved from a fossorial (burrowing) lizard,
or an aquatic lizard is finally solved. Snake osteology is very fragile and is
not a good candidate for fossilizing. But with new fossil finds and DNA
sequencing, the riddle has been solved and I am glad to say it was the route I
always wagered on and that is...snakes evolved from burrowing proto-lizard
varanoids, not just once but multiple times.
 |
| Snake evolution |
Snakes inherited the ability to produce
venom as most extant platynoans do in various volumes. Venom is primarily derived
proteins whereas snake venom evolved a single time from its lizard lineage
before divergence in speciation. The derived venomous protein further evolved
in snakes through gene encoding of a normal protein from regions of other
regulatory bioactivity processes. For example one pancreatic protein would be
recruited into a venomous proteome (a genome expressed entire set of proteins)
while protein from another body area is inducted also into the venom gland.
 |
| Venom clade |
This protein mixture eventually resulted
in a species unique venomous cocktail specific for affecting and subduing a
particular prey. Venom was developed for apprehending prey. Its secondary
advantage is in defense.
There may be no real way to know if
mosasaurs had the ability to inject venom. Although they came from a
terrestrial ancestor with venom injecting capabilities, the type mode of
injection is not from a hollow fang, as is the case for most recent evolved venomous
snakes, but from grooved fangs that require chewing to stimulate venom to flow
down the grooves and into the wound. In an aquatic environment mosasaur venom
would simply have washed away.
So perhaps, mosasaurs lost venom
injection capabilities, but it was no real loss as their jaws and size would
eventually allow them to rule the seas.
The most primitive extant basal snake
group is scolecophidians (blind snakes). One of the largest genetic reptile
dataset assemblages carried out by Associate Professor, John J. Wiens in the
Department of Ecology/Evolution at ‘Stony Brook University’ verifies almost all
extant snake groups arose from this primitive burrowing blind snake group. This
is highly suggestive in alluding to the fact that snakes had fossorial origins and
from this evolvement, all extant snakes still carry (no matter if they are
terrestrial, arboreal or aquatic) a retained small tail. Small tails are
evident in all burrowing fossorial vertebrate animals.
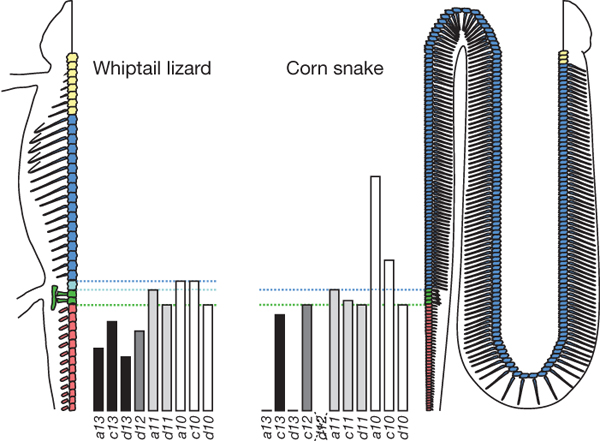
Hox genes govern the boundaries of the
neck, trunk, lumbar, sacral and tail regions of vertebrates. Hox genes play the
role of a switch-box controlling the modules of genes at specific regions along
the axis of the body. Hox genes encode a class of transcription factors and are
pivotal in specifying regional identity in body plans. Differences in their
expression explain the evolution of animal phyla. For vertebrates, Hox genes
function to regulate the regionalization of the axial skeletal structure. In
snakes, Hox genes have not turned off but have slowed the development of limb
structure, while greatly slowing it down in the forelimbs from the scapula (shoulder)
down to the phalanges. The slowing process has continually occurred until in
modern snakes, there is no evidence left of the forelimbs and only vestigial
remnants of the hind pelvic girdle in the more primitive snakes such as the
blind snakes and boas. This is totally the opposite effect in mammalian Hox
gene expression where the trending has been accelerated in limb lengths.
 |
| Add caption |
With recent fossil finds, the Hox gene
limb development slowdown has been nicely presented while pushing snake
evolution back another 70 million years from some 97 mya to 167 mya. Between
167-100 million years, snakes began radiating and evolving under their
increasing elongate and loss of limb body plans. In leaving a fossorial lifestyle and going back
to the surface, loss of limbs was an advantage for snakes in rocky/bushy
terrain, climbing and in swimming. Also
expressed in snake evolution is a reversion back to sight; although it isn’t
the most dependent sense snakes rely on.
Evolution of the elongated cylindrical
snake body reveals homoplasy in amniote Hox gene function. What this means is
that the snake body form is convergent being a shared characteristic between
differing species and not acquired from a common ancestor. Regionalized
expressions of the Hox gene code in the fossil record supports a subtle
morphological gradient along the anterior to posterior primaxial (along or
closer to) axis in stem member species of extant lizards and snakes.
So, even though the lack of Hox gene
expression in creating smaller and smaller limbs is apparent, the osteological
origin of the whole snake form is due more from the decoupling of primaxial and
abaxial (situated away from axis) domains in conjunction with increased somite
numbers. Therefore, primaxial Hox gene function does not play a part in the
overall snake skeletal structure.
Paleontologists now realize that the snake
overall body form was not due simply to the lowering of Hox gene expression,
for if that was so, then there would be fewer regional differences in the
shapes of vertebra down the vertebrae column. As it turns out, snakes have the
exact same number of differing vertebrae regions as lizards possess.
 |
| Eophis |

 |
| Diablophis |

The stem-snake genus, Parviraptor (phonectics: Par-vi-rap-tor) now includes just one fossil
species in P. estesi found in the very
late Jurassic to very Early Cretaceous 147-143mya England limestone bedrock. P. gilmorei, was initially designated in
the genus Parviraptor, but under more
phylogenetic scrutiny has been placed into the new genus Diablophis as explained above. Parviraptor
appears to have been restricted to lagoonal shallow subtidal swampy marine
environments.
These four snake finds fills a
morphological, phylogenetic and ecological data gap that was predicted by
molecular phylogenetics. It occurred during the final Pangaea breakup into Laurasia
and Gondwana co-occurring with early anguimorph lizards. The finds also bear
out that the evolution of characteristic snake skull elements appeared long
before the loss of limbs.
 |
| Ophidia clade |
Ophidia (phonetics:
O-fid-ee-ah) is a suborder for all extinct and extant snakes. The cladogram
below gives perspective in relationships to the ancient stem snakes or
proto-snakes to extant species.
 |
| Tetrapodophis fossil |
 |
| Tetrapodophis |
Found in northeastern Brazil’s
Cretaceous fossil rich limestone Crato Formation, Najash rionegrina (phonetics: Nah-jas Rye-un-eg-ree-nah)
proto-snake had lost its forelimbs, but had functional forelimbs outside the
ribcage including a well-defined sacrum supporting a pelvis. It was found in 90
million year old terrestrial deposits in Patagonia Argentina. Measuring up to 1.52m/5ft,
it was a burrowing snake.
 |
| Najash |
All three of these snakes’ have left fossilized paddle-like tail fin impressions much like modern seas snakes. In addition, the 1m/3.3ft long Pachyrhachis had very thick and dense ribs and vertebrae aiding it in diving.
 |
| Lft: Eupodophis Middle: Pachyrhachis Rt: Haasiophis |
| Lft: coniophis Rt: extant snake |
 |
| Snake cranial kinesis evolution |
Coniophis was a transitional snake in showing the evolution of snake cranial kinesis. In considering snake skulls, it represents the most primitive snake line, but is not the oldest snake. In the clade above, each circled number indicates cranial kinesis evolution with 1 representing the most primitive ascending to 10 as the most advanced form.
 |
| Titanoboa |
 |
| Superimposed Titanoboa skeleton |
Titanoboa (Phonetics:
Ty-tan-o-bo-ah) was a monster among predators reaching a length of 14.6m/48ft
and weighing in at 1,134kg/2,500lb. This boid occurred 60-58 mya in the
Paleocene just after the extinction of dinosaurs at the end of the Cretaceous.
With multiple remains found (28 individuals) at the Cerrejon coal mines of
Columbia, this snake lived in a warm tropical wet environment. although due to
the size of Titanoboa, the
environment could not have been too warm, for the giant snake’s metabolism
generated a lot of heat. Too hot a climate, the snake would’ve overheated and
perished. however, the warm temperatures did allow for the larger size as it
does today. Large boids (boas and pythons) today live in the tropics where the
smaller snakes inhabit more temperate climes.
Although Titanoboa could easily consume the largest crocodilians present at
the time, evidence alludes to the fact that the snake was piscivorous in
primarily preying on fish. This snake
was much thicker in its midsection than in the more tapered anterior and
posterior ends and most likely was a drab color for better camouflage as it was
an ambush predator.
 |
| An adult L. carlae |
Now compare the 14.6m/48ft Titanoboa to the extant snake, Leptotyphlops carlae (Phonetics:
Lep-toe-ty-flops Car-lay) which could comfortably curl up on a U.S. quarter at
10cm/3.9in.
Amphisbaenians:
Snakes and amphisbaenians evolved from
burrowing lizards. Both had fossorial origins, but where snakes went back to
the surface away from the tunneling and burrowing underground lifestyle,
amphisbaenians continued to exist in a fossorial environment. Instead of
primarily invertebrate prey found in tunneling, snakes found larger prey on
land, so the skull elements became more flexible and unhinged, where
amphisbaenians remaining underground retained more fused skull features to aid
in tunneling.
 |
| Amphisbaenian clade |
Amphisbaenians have been very difficult
to quantify, for although they indeed came from a lacertid lizard ancestry,
there too occurred much parallelism and convergent evolution between the two
groups having nothing to do with genetic sharing.
While amphisbaenians are mostly
limbless, three extant Bipes
(Phonetices: By-pees or Bi-pez) species have reduced forms of front limbs and
rudimentary internal hind limbs. Morphological data shows that species with
front limbs form a sister group to those that are limbless. This means that the
amphisbaenian loss of limbs occurred only once.
Tamaulipasaurus (phonetics: Tah-moe-lee-pa-sor-us)
was indeed a burrowing lepidosaur diapsid and also has numerous sister taxa.
Occurring in the Early Jurassic 189.6-183 mya and Middle Jurassic 165mya, two
fossils have been found in Mexico’s La Boca formation material. Extant
amphisbaenians and ambamids are closely related. One of Tamaulipasaurus’ sister groups gave rise to amphisbaenians while
another to dibamids.
 |
| Tamaulipasaurus fossilized skull |
Named Crythiosaurus (phonetics: Crith-ee-o-sore-us) this ~90 million year old lizard may well be a basal predecessor to amphisbaenians. Only the skull has been found and is longer and narrower with a larger occipital as is the case in lacertid lizards. However, extensive cranial fusion of skull elements are evident with the temporal reduced to just a piece of bone. Even though the skull was more elongate and narrower like lizards, the fusing of the skull elements suggest a fossorial lifestyle and it most likely had reduced limbs and an elongated body for burrowing. The teeth indicate an insectivorous diet.
Chalcides (Phonetics:
Chale-side) is an ancient and extant three-toed skink genus where embryological
evidence points to the lessening size and number of digits are due to Hox gene
d11 expressing digits 1, 2 and 3 as digits 2, 3 and 4. This suggests that digit
reduction, as in amphisbaenians, is a more frequent consequence in convergent
evolution such as in snake, dibamid lizards and avian evolution.
From an ancient sister group to Chalcides derived the genus, Sineoamphisbaena, which, although there
is strong debate, is one of the first stem amphisbaenians. This animal’s skull reveals
a mosaic of lizard and amphisbaenian like characters. A main feature is cranial
consolidation adapted for a fossorial lifestyle. However, the fossils show
skink-like modifications of the palate and temporal regions of the skull. The
relative primitive morphology of Sineoamphisbaena
is an indication that it did not devote its whole life to being fossorial and
most likely frequented the surface far more often than extant amphisbaenians
do. This most likely is due to the fact that during the animal’s time in the Late
Cretaceous, there was a transition from a perennial lacustrine environment to a
semiarid aeolian environment 75-71 mya in what is now present day Mongolia.
 |
| Skull comparison |
 |
| Sineoamphisbaena |
Sineoamphisbaena caused a
quandary for paleontologists, for the assumed basal and dispersion of
amphisbaenians was rooted in North America from 22 assigned Rhineura (Phonetics: Ree-noor-ah) species
and not from Inner Mongolia. From the 22 species, they were whittled down to 9
extinct and the only extant N. American in the species R. floridana. The debate still lingers, but Rhineura stills holds onto ground when considering extant
amphisbaenians. We’ll touch on that in a bit.
Cryptolacerta (phonetics:
Crip-toe-lah-sir-tah) appearing 47 mya is considered a lacertibaean as it had
an amphisbaenian fused head but, though much smaller, lacertid-like limbs. This
fossil, as a holotype supports molecular genetic studies that amphisbaenians
and lacertids are related.
After X-ray computed tomography analysis
on the Cryptolacerta well preserved
almost complete fossil with only the tip of the tail missing, results bear out
that the reinforced thickened skull is in a relationship with amphisbaenians
and that both are related to lacertids. This prognosis reveals the fact that
amphisbaenians’ cranial akinesis (skull element fusion) came first before the
snake-like body and loss of limbs during their evolvement. This also reveals
why amphisbaenians adapted to tunneling with the head.
 |
| Cryptolacerta fossil |
Cryptolacerta, through
studies with extant lizards present the lizard as an animal that primarily
spent its time with its body held low to the ground (squat bodied) while
leafing through forest floor litter, but was also an opportunistic burrower.
Cryptolacerta sits at the
base of amphisbaenian branch with amphisbaenians and lacertids branching out
from that base on differing, but adjacent lines.
 |
| Cryptolacerta |
In representing the family, Blanidae (phonetics: Blan-ah-day) is the
Miocene Epoch 11.6 mya fossil, Blanus
mendezi (phonetics: Blan-us
Men-de-zee) excavated in Valles-Penedes Basin in Spain’s Catalonia region.
Extant blanids are only represented in Europe, but further subdivided as western
and eastern, while as a whole, is one of the three groups representative of the
Mediterranean amphisbaenian groups.
 |
| B. mendezi skull |
The fossil is of a complete 5.8mm/0.23in
tiny skull encased in carbonate. Using computed tomography (CT), the scientists
developed a 3-D image showing it had 20 teeth and the skull was similar to
modern day blanids.
Of the three Mediterranean groups, one is
Iberian (western), another is Eastern Europe and the last is Northwest Africa.
This fossil represents the western group and alludes to detail that the Iberian
and Northwest African groups arose from one western Mediterranean group that
only later subdivided.
Currently, there are six families of
amphisbaenians and they are:
1)
Amphisbaenidae -
Amphisbaenids; tropical worm lizards of South America, some Caribbean islands,
and Sub-Saharan Africa (17 genera)
2)
Bipedidae - Only in
Mexico; commonly called ajolotes (1 genus)
3)
Blanidae - Anatolian,
Iberian, and Moroccan worm lizards (1 genus)
4)
Cadeidae - Cuban keel-headed
worm lizards; traditionally amphisbaenids, but shown by DNA to be closest to
Blanidae (1 genus)
5)
Trogonophidae - Palearctic
worm lizards (4 genera)
6)
Rhineuridae - North
American worm lizards (1 genus)
There are four distinct cranial
assemblages and all are highly specialized for a style of utilizing the head as
a tool for tunneling. These four modes of tunneling are associated with an
inter species stereotyped burrowing behavior.
 |
| A basal burrowing amphisbaenian fossil |
Bipedids and blanids use their blunt
‘round-headed’ form for tunneling by ramming the head into the soil and
widening the tunnel by uplifting the head and compacting the dug soil to the
tunnel ceiling. Bipedids also use their forelimbs to propel forward.
Rhineurids use a depressed ‘shovel-headed’
form by using the dorsoventrally flattened snout and with a strong craniofacial
angle, push the loose dirt to the sides where it is then compacted to the walls
of the tunnel as the body goes through.
Trogonophids use a ‘spade-headed’ form
by also using a dorsoventrally flattened snout, but with a strong craniofacial
angle and further use their lateralcanthi (sharp edging on sides of head) to
shave off soil from the front of the tunnel in an oscillatory fashion then pack
it to the tunnel sides with sides of the head and body.
Most amphisbaenids (8 genera) and the
cadeid utilize a compressed ‘keeled-headed’ form and dig with a laterally
compressed head by ramming the head forwards then push and pack the loosened
soil rearwards in pushing the head in opposite directions to pack the soil to
the floor and ceiling of the tunnel.
Due to the numerous convergent
evolutions within the amphisbaenian group, limited outgroup morphologies and
the lack of findings of the tiny and frail subterranean fossils, amphisbaenian
evolution has been a booger-bear to track and sort out for scientists.
 |
| Extant Amphisbaenian cladeogram |
Another quandary concerning amphisbaenians
is biogeography in how they became so widespread in living such a fossorial
lifestyle, for they certainly didn’t tunnel across the oceans. Amphisbaenians’
global dispersal began around 65 mya. They may have traveled on the third and
final breakup of Pangaea and Laurasia ending 60-55 mya, but that breakup began
a good 95 mya. What scientists feel is that they ocean rafted amongst soils
held together by roots. This also explains why other fossorial species are so
widespread.
Anyway, some primitive amphisbaenians
did make it to North America and diversified coming up with nine species of
rhineurids 60 mya. Eight species went extinct. The extant R. floridana is the only survivor because it was the first to
diverge and migrate where milder North American temperatures prevailed. The
extinct species’ fossils are found in the northern portion of N. America in
strata laid out during the Cenozoic’s Paleocene and Eocene Epochs 66-33.9 mya.
The climate was very mild then; there were even crocodiles swimming off the
coast of Greenland. By the beginning of the Oligocene 33.8 mya, cooler
temperate climates began to prevail while the Antarctic got its first ice cap.
The cold was too much for the eight warmer weather loving rhineurids. Not
enough cold reached into R. floridana’s
more southern range.
 |
| In my opinion molecular takes precedence over morphology |
For living amphisbaenians, as bipedids
are a sister taxon to the trogonophid-amphisbaenid clade, R. floridana as a Rhinenuridae
family member is the most basal morphological
species among other amphisbaenian families, but is a young genus first arriving
in the Pleistocene.
Euryapsida:
The first diapsid to experiment with
euryapsid skull fenestra was Araeoscelis
(phonetics: Ah-ray-oss-kell-iss) living 284.4-275.6 mya. This 60cm/2ft long reptile,
most likely gave up on its lower fenestrae to create a stronger bite, but it
had no direct basal relations to the marine euryapsids.
Euryapsids are lepidosaur reptiles
derived from diapsids in migrating the two single openings on each side into
the upper temporal as an opening behind each eye on top of the head. Euryapsids
had the diapsid lower temporal fenestrae fused over. Ichthyosaurs furthered the
euryapsid arrangement with a special set of introduced bones along the margins
of the fenestra (opening).
The euryapsid group is considered
polyphyletic in that most of their shared characteristics are considered
convergent evolution rather than related through ancestral genetics. They first
appeared in the early Triassic 245 mya and became extinct at the end of the
Cretaceous 65 mya.
 |
| Araeoscelis |
 |
| Araeoscelis basal clade |
There are some that still contend
euryapsids were archosaurs instead of lepidosaurs, but from the shoulder
girdles to the phalanges, sauropterygian limb anatomies and the lateral
undulation of the vertebral column, euryapsids were of lepidosaur origin. The
major euryapsid groups are divided into two major taxon superorders; they are:
 |
| Sauropterygian clade |
Sauropterygia (phonetics:
Sawr-op-ter-ridge-ee-ah): meaning ‘lizard flippers’ including the four orders, 1.
Placodontia (placodonts) 2. Pachypleurosauria (pachypleurosaurs)
3. Nothosauroidea
(nothosaurs) 4. Pleisosauria (the suborder:
plesiosaurs and suborder: pliosaurs).
 |
| Ichthyosauria clade |
Ichthyopterygia (phonetics:
Ick-the-op-ter-ig-ee-ah): meaning ‘fish flippers’ includes the two orders, 1. Grippidea (grippiosaur, gulosaur and
chaohusaur) 2. Ichthyosauria
(ichthyosaurs).
In sauropterygian evolvement, the major
ancestral suspect is the Late Permian neodiapsid belonging to the superfamily, Younginiformes (phonetics:
Yun-gin-nee-forms) that also survived into the Early Triassic. It is from the
tangasaurid family and further divided into the subfamily, Tangasaurinae (phonetics: Tain-gah-sor-ee-nay). While the other
tangasaurid subfamily, Kenyasaurinae
(phonetics: Ken-ya-sor-ee-nay) was terrestrial, Tangasaurinae species had become aquatic in a lacustrine
(freshwater or salty lake) environment. Adapted to an aquatic life with highly
derived amphibious characteristics, they had webbed feet and laterally
compressed tails for sinusoidal propulsion through water.
As mentioned earlier under diapsids, the
neodiapsid claudiosaurs are perhaps basal to nothosaurs, plesiosaurs and
pliosaurs. Considered an extension of the condition seen in claudiosaurs, pachypleurosaurs
and nothosaur palates are posterior and medial extensions of the pterygoids,
with the interpterygoid vacuities completely closed while ventrally, the base
of the braincase is covered.
 |
| Tangasaurid Hovasaurus |
| Claudiosaur |

Primitive sauropterygians like the
placodonts first appear in the Middle Triassic 245 mya and begin radiating quickly
just 5 million years after the great Permian/Triassic mass extinction (PTME).
Normally the bodies were squat and bulky with short legs and fed along shallow
coastal sea beds. By the Late Triassic 210 mya, placodonts had become extinct.
Placodonts experienced pachyostosis
giving heavier and denser bones for negative buoyancy to aid in being anchored
while feeding on shallow coastal sea beds. This still tied placodonts to land to
rest after both long periods of holding the breath in feeding and extraneous
swimming.
 |
| Placodonts |
 |
| Pachypleurosaur |
By 242 mya, pachypleurosaurs made their
appearance. These primitive sauropterygians, although small ranging in size
from 20cm/8in to 1m/3.3ft appear to be ancestral to plesiosaurs with small
heads, long necks and paddle-like limbs. Without doubt though, pachypleurosaurs
are a sister taxon to the eusauropterygian clade that both nothosaurs and
plesiosaurs belong in.
 |
| Nothosaur clade |
Soon after the appearance of
pachypleurosaurs, eusauropterygian nothosaurs appeared showing a rapid euryapsid
radiation and diversification in shallow Triassic epicontinental seas and near-shore
basins of the Tethyan seashore fringes. Once flooding from sea transgressions
in what is now the continent of Europe, eusauropterygians successfully followed
behind the sea inundation filling in the ecological niches as apex predators
from what is now Spain to China.
 |
| Nothosaur |
Nothosaurs were much like the present
day pinnipeds (seals, sea lions, walruses). Dependent upon the sea for much of
their food, but would clamor to the seashore for rest and sunbathing.
By the time nothosaurs died out,
plesiosaurs began to appear 203.6 mya in the Late Triassic. There is good
argument that nothosaurs evolved into plesiosaurs. The intermediate between
this nothosaur/plesiosaur transition was quite possibly the related pistosaurids.
Resembling nothosaurs, but more derived for a permanent aquatic life with
webbed feet replaced by flippers, they also had other plesiosaur features. Like
a nothosaur, pistosaurids had the same body form and the same primitive palate
with a mouth possessing the same type numerous sharp teeth. Like the
plesiosaur, it had flippers for maneuvering through water, a stiff vertebral
column and a similar skull structure and head.
 |
| pistosaur |
 |
| Plesiosuar/Pliosaur clades |
Plesiosaurs started off with short necks
in the earlier species much like nothosaurs. During the earliest part of the
Jurassic, a plesiosaur family subgroup, Rhomaleosauridae
(phonetics: Row-may-lay-oh-sor-ah-day) had developed into numerous species and
proves to be a basal split-off of short necked plesiosaurs. From these
subgroups towards the end of the Early Jurassic 180 mya, the long necked
plesiosaurs begin to appear.
Neck lengths continued growing until apexing with Elasmosaurus (phonetics: E-las-mo-saw-rus). Evolving and living in the Late Cretaceous 83.6-80.5 mya its neck at, 7.6m/25ft was at least half its 15.2m/50ft total length. Longer necks allowed for prey ambush strategies separating its visually bulky body from its mouth and supported a head with excellent eye vision in water, but watery environments totally supported the neck as it would not have been able to hold it above water.
 |
| Rhomaleosaurid |
 |
| Early plesiosaur |
Neck lengths continued growing until apexing with Elasmosaurus (phonetics: E-las-mo-saw-rus). Evolving and living in the Late Cretaceous 83.6-80.5 mya its neck at, 7.6m/25ft was at least half its 15.2m/50ft total length. Longer necks allowed for prey ambush strategies separating its visually bulky body from its mouth and supported a head with excellent eye vision in water, but watery environments totally supported the neck as it would not have been able to hold it above water.
 |
| Elasmosaurus |
 |
| Albertonectes |
 |
| Plesiosaur-Pliosaur neck comparison |
 |
| Liopleurodon |
 |
| P. funkei biting force 4x T. rex |
 |
| Pliosaurus funkei |
The first ichthyosaurs looked more like tuna while the latter species appeared more dolphin-like. In either form, all ichthyosaurs were streamlined for swiftness.
 |
| Thaisaurus |
Occurring in the Early Triassic so far, Thaisaurus (phonetics: Tie-saw-rus) is
the most basal ichthyosaur. Although the forelimbs had become flippers, the
hind limbs had not still exposing the feet phalanges and claws. Two other, but
more evolved ichthyosaurs showing up a bit later in the Early Triassic were Grippia (phonetics: Grip-pee-ah) and Utatsusaurus (phonetics: U-taht-sue-saw-rus).
These two had all four limbs entombed as flippers but did not yet support a
dorsal fin as later species would.
 |
| S. sikanniensis |
Still without a dorsal fin, appearing in
the Late Triassic 215-210 mya, Shastasaurus
sikanniensis (phonetics: Shas-tah-saw-rus
See-kan-nee-n-sis) is the largest marine reptile known growing to lengths of
21m/69ft. How it got that large is still a mystery, for all the species in the
genus Shasatsaurus possessed no teeth
and it is not known on what it ate or how it captured and ate its meals. Decent
speculation is that it grabbed and ate soft bodied cephalopods, but if you
truly come up with the answer you will be called a very clever person by many a
paleontologist.
 |
| Shastasaurus species |
Mixosaurus (phonetics:
Mis-o-saw-rus) of the Middle Triassic 230 mya has had its fossils found all
over the world. As the name implies it was a transitional ichthyosaur from the
early smaller and ungainly ones to the later more streamlined. Mixosaurs were
also one of the first ichthyosaurs to have a dorsal fin.
 |
| Mixosaurus |
 |
| Largest eye |

Although fossil eggs or fetuses of
placodonts have not been found yet, the evidence from all other euryapsid
fossil finds of embryos, fetuses, births and neonates prove most euryapsids gave
live births.
A recent ichthyosaur fossil find of a Chaohusaurus (phonetics: Kay-uh-hu-saw-rus)
mother dying while giving birth shows the infant ichthyosaur exiting head
first. There were three more fetuses also lined up head first to exit. The
reason for the death of the mother and her newborns was due to labor
complications. The main significance of this fossil find is that the birth was
head first.
 |
| Chaohusaurus birth |
 |
| Later evolved tail first births |
All tetrapodal marine life (whether
extinct or extant) that first derived from a land air breathing lineage then
had a latter ancestry of going back to the seas gives live birth in water as
tail first to avoid drowning of the newborn. There are innumerable fossil finds
of tail first euryapsid births. All extinct marine tetrapods (mosasaurs,
plesiosaurs, nothosaurs, etc.) and all extant marine tetrapods (whales,
dolphins, porpoises, etc.) give tail first births. This is to allow the maximum
time in being able to reach the surface for that first breath of air. When the
watery environment hits the head, breathing must occur. That’s why tail first is
an advantage by allowing the head to be the last sequence of the birth, giving
more time to reach that required first surface breath.
All land animals give birth head first
for the same reason, but as an opposite effect. Once the marine newborn is out
it must reach the surface quickly, so tail first is an advantage. But in land
animal births that breathe air, it is not. The infant needs instant access to air while
being born to keep it from suffocating while still in the birth canal during
delivery.
Both strategies have the same effect, to
reach air, but with opposite timing; maximum time for a tail first water birth
and minimum time for a head first atmospheric birth. What the more primitive
Early Triassic Chaohusaurus
implication states is that live birth for euryapsids first arose on land then
was adapted to watery environs evolving from head first to tail first over a certain
period of time.
 |
| Ichthyosaurus |
The video below is an example of how
most likely an ichthyosaur birth occurred.
Before we depart, let’s check out a
marine euryapsid living dilemma they had to overcome before being as successful
as they were.
Fish have no problem maintaining a high
level of metabolic energy consumption during locomotory paced activity, for external
gills give all the respiratory oxygen exchange needed without being hindered by
the fish’s flexing side-to-side movement during mobility.
The first tetrapods to walk on land
inherited that flexible side-to-side body motion to travel, but now, in converting
air bladders into internal lung respirators replacing gills, an interrupted
breathing dilemma arose. This inherited side-flex fish trait equated into a
sprawling gait and with the side-to-side body motion in locomotion caused a
problem for internal lung function. The first poor tetrapods walked two paces
then had to stop just to gasp for air in replenishing its oxygen supply.
Running was out of the question for early terrestrial travels.
Respiration and locomotion use the same
set of muscles as derived from fish ancestry. The trunk is flexed to the right
and left as land animals walk and run.
Although it may breathe when walking, it
cannot breathe when running. This is why.
As a tetrapod land animal steps forward
with left foot first, the right side of the thorax including the right lung is
compressed while simultaneously the left side of the thorax and left lung are
expanded. Of course the cycle reverses when the right foot moves forward.
So this is a big problem, for land
animals breathe in air to fill both lungs simultaneously, then exhale to
deflate the lungs for another breath of fresh air. But breathing normally
cannot commence when one lung is compressed. This distortion interferes with
normal thoracic breathing.
When walking slowly, the tetrapod may be
able to breathe between steps because the thorax has time to recover from the
distortion. But if the animal speeds up, the animal cannot breathe when
running. That is why tetrapods must stop in short periods to simply breathe
properly. This is called ‘Carriers Constraint’.
Land vertebrates have solved Carriers Constraint, especially mammals, by evolving a more erect stance with all four limbs vertically supporting the body. This aided moving in a vertical plane when running resulting in less twisting of the thorax, so sparse breathing could be possible during rapid locomotion. With bipedal motion as in humans and kangaroos, the thorax is lifted perpendicular to the ground allowing the ribcage to not flex during rapid locomotion. Still though, a breath can only be taken every other stride or hop.
 |
| Carriers Constraint lung compression |
Land vertebrates have solved Carriers Constraint, especially mammals, by evolving a more erect stance with all four limbs vertically supporting the body. This aided moving in a vertical plane when running resulting in less twisting of the thorax, so sparse breathing could be possible during rapid locomotion. With bipedal motion as in humans and kangaroos, the thorax is lifted perpendicular to the ground allowing the ribcage to not flex during rapid locomotion. Still though, a breath can only be taken every other stride or hop.
However a bipedal posture is
intrinsically unstable for it requires long periods of high level energy and
its consumption, so a high metabolic rate must be maintained at all times, even
during rest.
Both quadruped and biped land animals
can run for a while without breathing in switching to anaerobic glycolysis. A
sprinter can run a 100 meter dash without breathing by switching to anaerobic
glycolysis, but this can only be maintained for a few seconds before further
circumventing oxygen debt and introducing dangerous levels of lactic acid in
the blood in burning lactose instead of oxygen.
Now, with marine reptiles equipped with
land heredities like lungs and limbs for mobility, how did they resolve
Carriers Constraint in going back to aquatic environs?
These lung possessing marine reptiles
competed quite well with gill possessing fish through various choices in
swimming styles that did not hinder normal functions of the thorax and lungs
within it. Marine reptiles must utilize one breath when diving or subsurface
swimming.
Although still dependent on frequenting
land, placodonts overcame the constraint in having a leisurely lifestyle going
after molluscs and crustaceans on the sea beds of shallow coastal waters.
Placodonts also were building epidermal armor that would have made the body
more rigid and stiffened the thorax enough to negate twisting from mobile
movement.
Pachypleurosaurs and basal pistosaurs
were swimming in undulatory anguilliform eel-like fashion without any
strengthening or hardening of the thorax. In this transitional mode, pachypleurosaurs
had not yet approached a Carriers Constraint solution, so were forced to remain
near shorelines.
The Nothosaurs, through a series of
species from earliest to most recent, developed larger forelimbs for paraxial locomotion
while stiffening the thorax and spine. This nixed undulatory propulsion in the
most advanced species utilizing the limbs and not the body for locomotion. The
modified limbs anchored to a strengthened ball-and-socket glenoid joint were
like hydrofoils negating the body as a necessary element for locomotion.
Plesiosaurs and pliosaurs used paraxial
locomotion in which synchronized breathing with surface propulsion. With
well-developed massive pelvic and pectoral girdles, all the propulsive force
was applied with the limbs directed away from the axis of motion which was of
course the body. Plesiosaurians were one of the first to fly through waters.
Ichthyosaurs have not yet fully revealed
their key to the unlocking of Carriers Constraint. At a glance in being
dolphin-like in appearance, one might think that ichthyosaurs were accomplished
long distance and rapid swimmers. Appearances can be deceiving. The torso was
not all that rigid and rib cages of ichthyosaurs were lightly built.
Major ichthyosaur propulsion came from
the tail that was anatomically decoupled from the body by a caudal peduncle.
They swam in an up and down undulating motion of the tail with a body that
doesn’t appear too rigid. With this ammunition, it appears like the ichthyosaur
was not prepared to do battle with the constraint.
Ichthyosaur body adaptations with an
undulating tail were geared for rapid acceleration and short bursts of rapid
speed, but no anatomy shows that it was geared for long distance swimming.
Unless, it leaped as dolphins and penguins currently do.
With ichthyosaur tail undulations for
rapid acceleration and a pectoral fin for upward force in propulsion, this
reptile may have leaped to gain an upper hand over Carriers Constraint.
Subsurface cruising expends energy due to waves just below the surface creating
a zone of drag. If ichthyosaurs leaped, it would’ve required much less
breathing and while in the air giving plenty of time to take one deep breath or
two short ones. The subsurface high drag zone could’ve been neutralized by
swimming through it at a high angle of 30 degrees or more which would be
conserving energy more so than simply maintaining swimming within the drag zone.
In
Continuum:
Indeed this was a long article, but it
could have been five times longer and still not cover all the bases. Hopefully
though there is enough information to remain fluid.
All of the fore, above and future ‘Et
Tunc’ series are not concrete fact, but are solidly based on fact. Information
derived has most other’s first hand researched input with some sprinkling and
dashes of my own stringing together output.
For the next ‘Et Tunc’ series we’ll be
discussing the archosaurs that led to crocodilians, pterosaurs, dinosaurs and
ultimately birds.
See ya then...
BJA
In Times Elapsed
02/27/2016
This comment has been removed by a blog administrator.
ReplyDeletehttps://saglamproxy.com
ReplyDeletemetin2 proxy
proxy satın al
knight online proxy
mobil proxy satın al
5EQQ8S